 2019, Vol. 37
2019, Vol. 37Institute of Oceanology, Chinese Academy of Sciences
Article Information
- LIU Qinghe, MA Lin, LI Xinzheng
- A new species of the genus Haloschizopera Lang, 1944 (Copepoda, Harpacticoida, Miraciidae, Diosaccinae) from the East China Sea
- Journal of Oceanology and Limnology, 37(5): 1698-1707
- http://dx.doi.org/10.1007/s00343-019-8361-0
Article History
- Received Dec. 18, 2018
- accepted in principle Feb. 20, 2019
- accepted for publication Feb. 25, 2019




2 University of Chinese Academy of Sciences, Beijing 100049, China;
3 Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China;
4 Laboratory for Marine Biology and Biotechnology, Pilot National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266237, China
The genus Haloschizopera Lang, 1944 belongs to the large family Miraciidae (subfamily Diosaccinae), which inhabits a wide range of sediment types from flocculent mud to coralline debris and often dominants the copepod fauna of sublittoral soft sediments (Moore, 1979; Chullasorn, 2009). Haloschizopera is currently composed of 21 species (Wells, 2007). Accordingly, species of the genus have a worldwide distribution, from Europe (H. pygmaea Norman and Sccot, 1905) to eastern Pacific (H. apprisea Gee and Fleeger, 1990), with a great variation in depth from the sublittoral zone (H. mathoi Monard, 1935) to abyssal environment (H. abyssi Becher, 1974). Because the taxonomic study of benthic copepods is carried out late in China, only a few records of benthic harpacticoids have been reported (Gee and Mu, 2000; Mu and Gee, 2000; Mu and Huys, 2002, 2004; Huys and Mu, 2008; Ma and Li, 2011, 2017a). Moreover, species of Miraciidae often occurs in ecological studies as a dominant group with high diversity or abundance (Mu et al., 2001; Ma and Li, 2017b). Therefore, the species of Miraciidae have great importance in both taxonomic and ecological studies.
During a biodiversity survey of benthic copepods in the sea area off the Changjiang (Yangtze) River estuary, several specimens of Haloschizopera were collected from the sublittoral zone. After a careful examination with other congeners, these specimens were confirmed to be a new species. Hence, it was described in the present study, representing the 22nd species of the genus.
2 MATERIAL AND METHODSamples were collected in Dec. 2016 near the offshore of Zhejiang Province in the East China Sea on R/V "Kexue-III". The sediments of the collection sites range from muddy sand to mud at a depth ranging from 25–79 m.
All the samples were fixed in 10% formalin and the specimens were extracted from benthic samples by a 31-μm sieve and colloidal silica Ludox HS-40 suspension centrifugation flotation method. Then the specimens were preserved in 75% alcohol. For their identification, the specimens were cleared in lactic acid and observed under a light microscope. Before dissection, the habitus was drawn and the whole-body length (from the basis of rostrum to the end of the anal somite) was measured while being temporarily mounted in lactophenol. Specimens were dissected in lactic acid, mounted on slides in lactophenol, and sealed with neutral balsam. Observations and dissection were operated with a stereomicroscope (Nikon C-DSD230), drawing by a microscope (Nikon ECLIPSE Ni-U) with the help of drawing tube. The habitus was drawn at 400× magnification; appendages were drawn at 1 000× magnification, with an oil immersion lens.
The terminology follows Huys et al. (1996). Abbreviations used in the text, figures and tables are: aes: aesthetasc; A1: antennule; L/W: length׃width; exp: exopod; exp (enp) -1(-2-3): the first (second, third) segment of exopod (endopod); P1–P6: swimming legs 1–6. All figures have been prepared with the aid of a drawing tube. The type material is deposited in the Marine Biological Museum, Chinese Academy of Sciences, Qingdao, China (MBMCAS).
3 RESULTThe systematics are shown below:
Order Harpacticoida Sars, 1903
Family Miraciidae Dana, 1846
Subfamily Diosaccinae Sars M., 1906
Genus Haloschizopera Lang, 1944
Haloschizopera sheni sp. nov.
3.1 Material examinedHolotype: 1 adult female, dissected on 8 slides (MBM189241), soft mud, depth 52.4 m, 122.508°E, 28.95°N, December 18, 2016. Paratypes: Three adult females, dissected on nine slides (MBM189242-189244); two adult males, dissected on six slides (MBM189245-189246); four adult females and four adult males (MBM189247) preserved in alcohol, collecting information as holotype.
3.2 EtymologyThe species is named after Dr. SHEN Chia-jui, in recognition of his numerous contributions to the freshwater and marine harpacticoid fauna of China.
3.3 DiagnosisBody subcylindrical, widest at the posterior margin of cephalosome, cephalothorax tapering anteriorly, cephalosome and thorax subequal, prosome slightly wider and longer than urosome. Rostrum defined at base, elongate, triangular, semi-hyaline distally, with central reinforcement, extending beyond second antennular segment, with one pair of small lateral sensilla in distal half-length. Antennule eight-segmented in the female, nine-segmented in male; aesthetascs on fourth and terminal segments. Allobasis of the antenna with one seta at inner middle margin; exopod three-segmented, second segment with one plumose seta; endopod with seven terminal elements. Mandibular basis with three plumose setae; exopod two-segmented, with 2׃3 setae; endopod one-segmented, with one middle and six terminal setae. Maxillary coxal endite with two setae; basis with six setae; endopod and exopod one-segmented with four naked setae and two plumose setae, respectively. Maxilla with three coxal endites, with 2׃2׃3 elements (proximal to posterior); allobasal endite with three elements; endopod with seven naked setae. Maxilliped with four setae on syncoxa; basis with two slender setae; endopod one-segmented, with a terminal claw and two accessory setae. P1 endopod and exopod three-segmented; endopod not prehensile, enp-1 extending not beyond the end of exp-2, sexual dimorphism characterized by one more spine and reinforcement at the inner margin of basis. P2-4 rami three-segmented, except enp-2 of P2 in male; P2 enp-2 modified with three outer setae, one apical spine and two inner spines in male, one of the inner spines swelled at tip; exp-3 of P2-4 with one, one, two inner setae, respectively. Caudal rami slightly shorter than broad, with one dorsal, two ventral, one inner lateral, one outer distal and two terminal setae.
3.4 Description of the femaleBody Length (from the base of the rostrum to the end of anal somite) 365–475 μm (mean=431 μm, n=6). Body subcylindrical (Fig. 1a), tapering slightly posteriorly, widest at the posterior margin of cephalosome. Rostrum defined at base, elongate, triangular, semi-hyaline distally, extending beyond second antennular segment, with central reinforcement and one pair of small lateral sensilla in distal half-length. Cephalothorax tapering anteriorly, cephalosome with scattered sensilla, thorax with paired sensilla; cephalosome and thorax subequal. Genital double-somite dorsally and laterally subdivided by a subcuticular rib; a genital area as in (Fig. 1c); vestigial sixth swimming leg bearing one long plumose seta and two short spines. Anal somite deeply divided dorsally with inner margins bearing complex rows of setules; operculum semicircular, near the median dorsal anterior border and overlain by posterior extension of a dorsal hyaline frill of penultimate somite. Caudal rami (Fig. 2) slightly shorter than broad (length/width: 0.82–0.88), with two spinules on inner margin; carrying seven setae, ventral surface with one distal spine located on small peduncle, and one adjacent slender seta inside distal inner margin, outer lateral surface with one naked and well-developed seta; inner subdistal corner with one small smooth seta; dorsal surface with one three-segmented seta; terminal margin with two well developed principal setae, strongly spinulose in median portion.

|
| Fig.1 Haloschizopera sheni sp. nov. a. habitus of adult ♀; b. lateral view of ♀; c. ventral view of ♀; d. lateral view of ♂; e. ventral view of ♂; f. habitus of adult ♂. |

|
| Fig.2 Haloschizopera sheni sp. nov. a. ventral surface of caudal ramus, ♀; b. dorsal surface of caudal ramus, ♀. |
Somatic ornamentation (Fig. 1a–c). Prosome with sensilla and pores. Urosome: dorsal surface smooth, with several pairs of sensilla except penultimate somites; genital double-somite with three and one dorsal-lateral rows of spinules on anterior part and posterior part, respectively; fourth urosomite with one dorsal-lateral row of spinules on posterior part; preanal somite with one lateral and one ventral rows of spinules at same level on posterior part; anal somite with lateroventral row of spinules around caudal rami. Posterior end margin of the cephalothorax and hyaline frills of urosomites striated, with dentate margin visible at 40× objective.
Antennule (Fig. 3a). Eight-segmented, first segment with setulose seta; second segment with large and small, posteriorly directed, unipinnate seta; distinctly unipinnate setae also on seventh and eighth segments; aesthetascs on fourth and terminal segments; armature: 1 [1], 2 [11], 3 [6], 4 [4 + aes], 5 [2], 6 [3], 7 [3], 8 [7 + aes].

|
| Fig.3 Haloschizopera sheni sp. nov. a. antennule, ♀; b. antennule, ♂; c. antenna; d. mandible; e. maxillule; f. maxilla; g. maxilliped; c–g: ♀. |
Antenna (Fig. 3c). Coxa with a row of setules. Allobasis with one pinnate seta at the proximal half of abexopodal margin. Exopod three-segmented, the first segment and second segment both with one plumose seta; second segment short; third segment slender, with spinules on outer margin, one plumose seta proximally, and two setae on distal margin, one pinnate and one naked. Endopod with two rows of setules on outer margin and distal margin; lateral armature consisting of several spines, one naked seta and two bifurcated setae; distal margin with seven elements: one pinnate seta, one slender seta, four geniculate setae, posterior one of which strongly spinulose around point of inflexion and sharing base with slender seta, one short fine seta inserted inside terminal edge.
Mandible (Fig. 3d). Gnathobase armed with one large tricuspid tooth, two bicuspids, four unicuspid teeth, and two smooth setae at outer distal corner. Basis broad, with three rows of spinules on the surface and three plumose setae on distal edge. Endopod one-segmented, with one naked lateral seta, three terminal setae, one of which pinnate, and three naked terminal setae fused at base. Exopod two-segmented, approximately half-length of endopod; a proximal segment with two plumose setae on the distal corner, the terminal segment with two plumose and one smooth seta.
Maxillule (Fig. 3e). Arthrite of praecoxa with two smooth setae on anterior surface; distal margin with six stouts and three slender spines. Coxa with two setae on distal margin, one stout, one slender. Basis bearing six elements, one well-developed seta on the subdistal corner, two subterminal setae, and three apical setae (one naked, one pinnate and one dentate). Exopod one-segmented, with two plumose setae. Endopod one-segmented, with four smooth apical setae.
Maxilla (Fig. 3f). Syncoxa with two rows of spinules at the outer proximal edge; inner edge with three endites, proximal endite armed with two spines, middle endite with one spine and one seta, distal endite with three spines (one pinnate). Allobasal endite with two spines (one dentate) and one naked seta fused at the base of endite. Endopod one-segmented, with seven smooth setae, of which one is stout and long.
Maxilliped (Fig. 3g). Syncoxa with one row of spinules on proximal edge; subdistal corner with two small pinnate setae; distal margin with two unequal pinnate setae. Basis with a row of spinules on the proximal surface; subdistal and middle inner margin each with one slender seta. Endopod one-segmented, with one terminal claw and two accessory setae.
P1 Intercoxal sclerite long, rectangular, with the upper margin swelled at the median (Fig. 4a). Coxa with two rows of spinules on anterior face. Basis with two rows of well-developed spinules at the proximal inner margin and distal outer corner; inner spine stout, strongly spinulose and with a row of spinules at base; outer spine pinnate. Exopod and endopod three-segmented, all segments with row of stout spinules along outer margin except exp-3; both exp-1 and -2 with one long pinnate spine at outer distal corner, and one row of setules on inner margin; exp-3 with two pinnate spines at outer distal corner and two geniculate apical plumose setae; enp-1 and -2 each with one plumose seta at subdistal inner margin; enp-3 with two apical setae (one large, geniculate, plumose and another small, smooth), and one pinnate spine at outer subdistal corner. Exopod shorter than Endopod, first and third segments subequal, slightly longer middle segment; Endopod not prehensile, enp-1 and -3 subequal, enp-2 shortest.

|
| Fig.4 Haloschizopera sheni sp. nov. a. P1, ♀; b. P2, ♀; c. basis of P1, ♂; d. endopod of P2, ♂. |
P2 Intercoxal sclerite bulbous, with two conical projections, unornamented (Fig. 4b). Coxa with two rows of spinules and one row of short setules on the anterior surface. Basis with one row of setules at inner distal corner, several minute setae at outer distal corner along with several spinules around the base and one spine at the outer proximal corner. Rami three-segmented, terminal segments longest; endopod slightly longer than exopod. The outer edge of enp-1 with setules, other segments with spinules. Enp-1 with the small sharp outer process at inner distal corner; enp-2 with the conspicuous sharp outer process at outer distal corner.
P3 (Fig. 5a). Intercoxal same as P2. Coxa with two rows of spinules and one row of short setules on the anterior surface. Basis with one row of setules at inner distal corner, several minute setae at outer distal corner along with several spinules around the base. Rami three-segmented, terminal segments longest; endopod subequal with exopod. The outer edge of all segments with spinules; a middle segment with the sharp outer process at outer distal corner.

|
| Fig.5 Haloschizopera sheni sp. nov. a. P3, ♀; b. P4, ♀; c. P3-exp-3, ♂. |
P4 (Fig. 5b). Intercoxal as P2. Coxa with two rows of spinules and one row of short setules on the anterior surface. Basis with one row of setules at inner distal corner, several minute setae at outer distal corner along with several spinules around the base and one spine at the outer proximal corner. Rami three-segmented, terminal segments longest; endopod shorter than exopod. The outer edge of enp-1 with setules, other segments with spinules; enp-2 with the conspicuous sharp outer process at outer distal corner.
P2–P4 setal formulae are as follows:
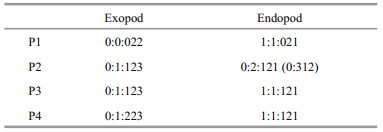
P5 (Fig. 6a). Baseoendopods not fused medially, separated with exopod. Baseoendopodal lobe slightly extending beyond half-length of exopod. Baseoendopod with one inner subterminal pinnate seta and three apical pinnate setae (middle one more than two times as long as others); outer margin with few spinules, the hyaline area near the base of exopod. Exopod slightly oval in shape, approximately 1.2–1.4 times as long as broad, with two rows of spinules on inner margin, and five setae (one inner, one apical, three outer); inner seta well developed and pinnate, about 4.1–4.2 times as long as apical seta, apical seta fine, naked, and borne on short peduncle; distal outer seta naked, as long as inner seta; middle outer seta pinnate, less than half length of distal outer seta; outermost seta small and naked.

|
| Fig.6 Haloschizopera sheni sp. nov. a. P5, ♀; b. P5, ♂. |
Similar to female except for the following features:
Body Shorter than females (325–367 μm). The first two abdominal somites are distinct.
Somatic ornamentation (Fig. 1d–f). Abdominal spinulation more complete than in female; first abdominal somite with one lateral and one dorsolateral row of spinules; second and third abdominal somites both with one continuous lateral-ventral row of spinules, one ventral row of short spinules on anterior part of second abdominal somite; fourth abdominal somite only with one continuous ventral row of spinules.
Antennule (Fig. 2b). Haplocer, nine-segmented, with major articulation between sixth and seventh segments; aesthetascs on fourth segment and terminal segment. Armature: 1 [1], 2 [10], 3 [7], 4 [3 + aes], 5 [1], 6 [1], 7 [1], 8[2], 9 [5+aes].
P1 (Fig. 4c). Basis with two chitinous projections near inner margin, one naked, another unipinnate.
P2 (Fig. 4d). Endopod two-segmented. The first segment ornamented with a row of setules on inner and outer margin. The second segment modified; proximal inner margin with one plumose seta, one small smooth seta arising from the small cuticular projection; distal margin with one strong inner plumose seta and one terminal spine; outer margin with stout spine and slender spine subterminally, slender spine swelled at the tip.
P3 (Fig. 5c). The distal segment of exopod with small hyaline tube pore issuing from the anterior surface, near the point of insertion of the inner seta.
P5 (Fig. 6b). Endopodal lobe extending beyond the end of exopod, with two stout pinnate terminal setae and few spinules along outer margin, hyaline spot at the base of exopod. Exopod separate, subcircular, with five setae; inner seta stout and pinnate; apical seta plumose, borne on small peduncle, ornamented with two spinules at base, longer than half length of inner seta; distal outer seta smooth, about twice as long as apical one; other two outer lateral setae small and naked.
P6 (Fig. 1e). Plate with one inner large seta and two small outer setae. One leg is larger than the other.
Variation No obvious variation was noticed within specimens except body length. Urosome and prosome often bent at an angle, which may lead the shrinking of urosomites and variation in length/width ratio of abdominal somites.
4 DISCUSSIONLang (1948) established the genus Haloschizopera with six species (including one synonym). Since then, another 16 valid species have been added into this genus. Species belonging to this genus usually small, semi-cylindrical with moderate abdominal ornamentation; A1 not elongate and with two strong plumose setae on the posterior margin of second segment; A2 with allobasis, exopod three-segmented and with one seta on second segment; mandible and maxillule biramous; maxilla and maxilliped uniramous, endopod one segmented; P1 enp-1 shorter than first two exopodal segments; number of inner setae on exp-3 of P2-4 reduced; P2 enp-2 with two inner setae; and in male, the modifications of P1 basis and P2 endopod, P3 exp-3 hyaline tube pore same. Thus, the characters shown by the new species suggest it should belong to the genus Haloschizopera.
Noticeably, we misidentified all the specimens as Sinamphiascus dominatus for the first time when examined by the stereomicroscope, as H. sheni is remarkably similar to S. dominatus Mu and Gee (2000) in habitus. However, the setal formulae of swimming legs and construction of P5 in both sexes of H. sheni differ greatly from that of S. dominatus: 1) in S. dominatus, exp-2 of P1, exp-1 and exp-3 of P2-P4 each with one more inner setae than that in H. sheni; exp-3 of P1 bearing one more minute seta on outer margin; 2) in S. dominatus, exopod of P5 bearing six setae and about twice as long as broad, endopod with five setae in female and two setae in male, while in H. sheni, exopod of P5 bearing only five setae and about 1.2 times as long as broad, endopod with four setae in female and two setae in male. In addition, we can still discriminate H. sheni from S. dominatus by the characters: proximal two segments of A1 thicker and broader, genital double-somite with three dorsal lateral rows of spinules on anterior part and fourth urosomite unornamented on the ventral surface.
The number of inner seta on exp-3 of P3 and P4 varies among the species within the genus Haloschizopera. Species may be categorized into four groups (Por, 1964; Becher, 1974) based on this character: (Group-A) P3 exp-3 with one inner seta and P4 exp-3 with two setae; (Group-B) P3 exp-3 without inner seta and P4 exp-3 with one inner seta; (Group-C) P3 exp-3 without inner seta and P4 exp-3 with two inner setae; (Group-D) P3 and P4 exp-3 both with one inner seta.
Haloschizopera sheni sp. nov. belongs to Group-A but it differs from the other species within this group significantly by the presence of one inner seta on P2 exp-3. In addition, we can still discriminate H. sheni from the other species within this group by the following features: P5 exopod about 1.2 times as long as wide (2 times in H. pygmaea Norman and Scott, 1905, 1.7 times in H. mathoi Monard, 1935, 3 times in H. marmarae Noodt, 1955 and 1.7 times in H. ruthorum Por, 1967); A1 eight-segmented (seven-segmented in H. bulbifera Sars, 1911, H. nuditerga Moore and O'Reilly, 1989, H. lionensis Moore and O'Reilly, 1989 and H. clotensis Moore and O'Reilly, 1989); P1 enp-1 extending not to end of exp-2 (H. ruthorum and H. tenuipes Noodt, 1964 both have long endopod of P1, P1 enp-1 extending to end of exp-2 in H. ruthorum, extending to end of exopod in H. tenuipes).
Species within the genus Haloschizopera rarely has inner seta on exp-3 of P2, except H. aegyptica Noodt, 1964. However, H. aegyptica has only one inner seta on P4 exp-3, belonging to the Group-D. In addition, exopod of P5 in the female is oblong, about two times as long as broad; P1 enp-1 extends to the middle point of exp-3; antennule is slender and prolong, ornamented with few armatures after the fourth segment in H. aegyptica.
5 CONCLUSIONA new species of the genus Haloschizopera from the East China Sea was identified. H. sheni sp. nov. can be easily identified by one inner seta on exp-3 of P2. According to its setal formulae, the new species belongs to the group with one inner seta and two inner setae on exp-3 of P3 and P4, respectively. H. sheni sp. nov. is very similar to Sinamphiascus dominates in habitus; therefore great caution must be paid when identifying these specimens.
6 DATA AVAILABILITY STATEMENTThe authors declare that all the data supporting the findings of this study are available within the article.
7 ACKNOWLEDGMENTWe thank Dr. MU Fanghong (Ocean University of China, Qingdao) for kindly providing valuable literature for our study on harpacticoida of the genus Haloschizopera. Many thanks also to Dr. KOU Qi of our team for his suggestions to improve the manuscript.
Becher K H. 1974. Eidonomie und taxonomie abyssaler harpacticoidea (crustacea, copepoda). Teil Ⅰ. Cerviniidaeameiridae. "Meteor"-Forsch-Ergebn D, 18: 1-28.
|
Chullasorn S. 2009. A review of Typhlamphiascus Lang, 1944(Copepoda:Harpacticoida:Miraciidae) with a new species Typhlamphiascus higginsi from Phuket Island, Thailand. Zoological Studies, 48(4): 493-507.
|
Gee J M, Fleeger J W. 1990. Haloschizopera apprisea, a new species of harpacticoid copepod from Alaska, and some observations on sexual dimorphism in the family Diosaccidae. Transactions of the American Microscopical Society, 109(3): 282-299.
DOI:10.2307/3226799 |
Gee J M, Mu F H. 2000. A new genus of Cletodidae (Copepoda; Harpacticoida) from the Bohai Sea, China. Journal of Natural History, 34(6): 809-822.
DOI:10.1080/002229300299273 |
Huys R, Gee J M, Moore C G, Hamond, R. 1996. Marine and brackish water harpacticoid copepods. Part 1. In: Synopses of the British Fauna. Field Studies Council, Shrewsbury. 352p.
|
Huys R, Mu F H. 2008. Description of a new species of Onychostenhelia Itô (Copepoda, Harpacticoida, Miraciidae) from the Bohai Sea, China. Zootaxa, 1706: 51-68.
DOI:10.11646/zootaxa.1706.1.2 |
Lang K. 1948. Monographie der Harpacticiden. Håkan Ohlsson, Lund, 2 vols. 1682p.
|
Ma L, Li X Z. 2011. Delavalia qingdaoensis sp. nov.(Harpacticoida, Miraciidae), a new copepod species from Jiaozhou Bay, Yellow Sea. Crustaceana, 84(9): 1085-1097.
DOI:10.1163/001121611X584334 |
Ma L, Li X Z. 2017a. A new species of the genus Typhlamphiascus (Copepoda, Harpacticoida, Miraciidae)from the South China Sea. Crustaceana, 90(7-10): 989-1004.
DOI:10.1163/15685403-00003679 |
Ma L, Li X Z. 2017b. Benthic harpacticoid copepods of Jiaozhou Bay, Qingdao. Chinese Journal of Oceanology and Limnology, 35(5): 1127-1133.
DOI:10.1007/s00343-017-6031-7 |
Monard A. 1935. Les harpacticoïdes marins de la région de Salammbo. Bulletin Station Oceanogrpahique de Salammbo, 34: 1-94.
|
Moore C G, O'Reilly M G. 1989. A re-examination of some problematical species of Haloschizopera (Copepoda, Harpacticoida). Journal of Natural History, 23(1): 93-110.
DOI:10.1080/00222938900770061 |
Moore C G. 1979. Analysis of the associations of meiobenthic Copepoda of the Irish Sea. Journal of the Marine Biological Association of the United Kingdom, 59(4): 831-849.
DOI:10.1017/S0025315400036870 |
Mu F H, Gee J M. 2000. Two new species of Bulbamphiascus(Copepoda:Harpacticoida:Diosaccidae) and a related new genus, from the Bohai Sea, China. Cahiers de Biologie Marine, 41(2): 103-135.
|
Mu F H, Huys R. 2002. New species of Stenhelia (Copepoda, Harpacticoida, Diosaccidae) from the Bohai Sea (China)with notes on subgeneric division and phylogenetic relationships. Cahiers de Biologie Marine, 43(2): 179-206.
|
Mu F H, Huys R. 2004. Canuellidae (Copepoda, Harpacticoida)from the Bohai Sea, China. Journal of Natural History, 38(1): 1-36.
DOI:10.1080/00222930210138935 |
Mu F H, Zhang Z N, Guo Y Q. 2001. The study on the community structure of benthic copepods in the Bohai Sea. Acta Oceanologica Sinica, 23(6): 120-127.
(in Chinese with English abstract) |
Noodt W. 1955. Marine Harpacticoiden (Crust. Cop.) aus dem Marmara Meer. Istanbul Universitesi fen Fakultesi Mecmuasi, 20: 49-94.
|
Noodt W. 1964. Copepoda Harpacticoidea aus dem Littoral des Roten Meeres. Kieler Meeresforschungen, 20: 128-154.
|
Norman A M, Scott T. 1905. Crustacea Copepoda new to science from Devon and Cornwall. Annals and Magazine of Natural History, 15: 284-300.
DOI:10.1080/03745480509443044 |
Por F D. 1964. Les Harpacticoides (Crustacea, Copepoda) des fonds meubles du Skagerak. Cahiers de Biologie Marine, 5(3): 233-270.
|
Por F D. 1967. Level bottom Harpacticoida (Crustacea, Copepoda) from Elat (Red Sea), Part Ⅰ. Israel Journal of Zoology, 16(3): 101-165.
|
Sars G O. 1911. An Account of the Crustacea of Norway.Volume V. Bergen Museum, Bergen. 449p.
|
Wells J B J. 2007. An Annotated Checklist and Keys to the Species of Copepoda Harpacticoida (Crustacea). Magnolia Press, Auckland. 872p.
|