 2017, Vol. 35
2017, Vol. 35Institute of Oceanology, Chinese Academy of Sciences
Article Information
- ZHANG Huajun(张华俊), PENG Liang(彭亮), GU Binhe(古滨河), HAN Bo-Ping(韩博平)
- Effect of trans-reservoir water supply on carbon and nitrogen stable isotope composition in hydrologically connected reservoirs in China
- Chinese Journal of Oceanology and Limnology, 35(5): 1117-1126
- http://dx.doi.org/10.1007/s00343-017-5327-y
Article History
- Received Nov. 20, 2015
- accepted in principle Feb. 2, 2016
- accepted for publication Jul. 27, 2016




2 Soil and Water Science Department, University of Florida, Gainesville, FL 32611, USA
Particulate organic matter (POM) is required to maintain the normal function of aquatic ecosystems and is typically derived from both autochthonous and allochthonous sources (Kling et al., 1992; Grey and Jones, 1999; Pace et al., 2004; Cole et al., 2011). In addition to phytoplankton, POM includes terrestrial sources such as soil organic matter and leaf debris, resuspended sediment, and in situ macrophyte and microzooplankton production (Gu and Schelske, 2010; Rožič et al., 2015). For an isolated lake or reservoir, POM usually shows a stable seasonality which depends on the relative importance of each component. Stable isotopes, especially of carbon and nitrogen (13C and 15N), are commonly used to investigate sources of organic matter and carbon and nitrogen biogeochemical processes in aquatic ecosystems (Kendall et al., 2001; Lehmann et al., 2004; Gu, 2009). Gu et al. (2011) suggested that the 13C in POM, 13CPOM, may also be an indicator of the relative importance of POM from in situ productivity and exogenous organic matter. Most investigations have focused on either a single lake (Yoshioka et al., 1994; Gu and Schelske, 1996; Jones et al., 1998) or a series of isolated lakes in a specific region (Grey et al., 2001; Gu, 2009; Hadas et al., 2009).
Reservoirs are highly hydrodynamic, usually with a shorter residence time than natural lakes, so in them, POM from exogenous organic matter often plays a relatively more important role. A number of recent studies used stable isotopes to track ecological processes on man-made reservoirs, which possess limnological features different from those of natural lakes (Shotbolt et al., 2005). For example, Hou et al. (2013a) investigated variations in δ13C and δ15N of POM in 20 reservoirs in southern China to evaluate the isotopes as indicators of primary productivity and trophic state. Primary productivity derived by high light intensity and temperature controls seasonality of 13CPOM, but δ15NPOM is mainly regulated by nutrient sources from pollution. Similarly, seasonal variations in POM and zooplankton δ13C and δ15N was examined to study the effects of highly variable inflows with the onset of the summer monsoon season in a deep reservoir in South Korea (Lee et al., 2013).
In tropical and subtropical coastal regions where freshwater is limited, many small or middle-sized reservoirs obtain pump water from nearby rivers and are then connected for effective water supply (Han and Liu, 2012; Zhang et al., 2013). In such cases, hydrological and biogeochemical characteristics of the water supplying reservoirs are expected to impact the receiving water body. Information about stable isotopes in POM allows us to compare similarity or dissimilarity of the composition of POM between connected water bodies, and even to recognize the particulates' mixed sources and transportation paths (Gu and Schelske, 2010; Gu et al., 2011).
The main purpose of this study is to investigate the seasonality of POM carbon and nitrogen stable isotopes in three hydrologically connected reservoirs and a river, and to determine to what extent the seasonal similarity for both δ13CPOM and δ15NPOM depends on connectivity between these tropical water bodies. In the present studied system, river water is first pumped into a reservoir (Dajingshan Reservoir, DJS), and then feeds a neighboring reservoir (Fenghuangshan Reservoir, FHS) via an underground culvert. A small reservoir (Meixi Reservoir, MEX) also feeds DJS but only in summer. As a result, the four water bodies form a gradient of connectivity.
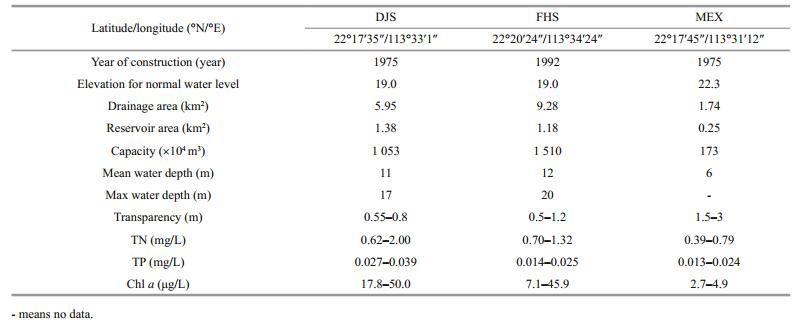
2 MATERIAL AND METHOD 2.1 Study areaThe three studied reservoirs, DJS, FHS and MEX and the water pumping station (Guangchang) are located in Zhuhai city, Guangdong Province, tropical China (21°48′–22°27′N, 113°03′–114°19′E) and downstream of the Zhujiang (Pearl) River basin. All reservoirs have relatively small catchments, and their water quality differs and largely depends on whether the receiving water is pumped or not (Table 1). The climate is of a typical monsoon nature with a wet season from April to September, followed by a dry season from October to March. The amount of precipitation from April to September accounts for about 85% of the entire year (Wen and Chen, 2005). The annual average air temperature in this region is 22.3℃. Although rich in rainfall (1 770–2 300 mm per year), Zhuhai has been challenged by water supply shortages because of an uneven temporal distribution (Wen and Chen, 2005), a coastal terrain, river pollution, and salt water intrusion. Thus, many reservoirs have been built to alleviate water shortages in the dry season (Han and Liu, 2012). Because reservoirs in Zhuhai are small to medium-sized with small catchment areas, they require water to be pumped from rivers to increase water storage. Water quality of the reservoirs is influenced directly by that of the source water from the rivers.
The three adjacent reservoirs of DJS, FHS and MEX are connected through a water transfer tunnel or a spillway. Water from Guangchang Pumping Station (GCPS) at Qianshan River, the end estuary of Zhujiang River is drawn into DJS (Fig. 1). It then flows from DJS into FHS via an underground culvert. MEX is the smallest of the three; its water overfills into DJS only during wet seasons for flood control. There are no farmlands or industries in the reservoirs' drainage basin, located in a mountainous area.

|
| Figure 1 Location of the three studied reservoirs and their connections to the GCPS in Zhuhai, the end estuary of Zhujiang River in Guangdong Province, China The arrow length indicates connectivity, i.e. the relative amount of the transported water. |
DJS with a surface area of 1.38 km2, was primarily built for farm irrigation in 1970s and was only used to supplement urban drinking water supply and flood control. Since 1980, its main function was altered to supply domestic water and to control flooding. It has become one of the important sources of water in the Zhuhai urban district. The annual average output for water supply is 2×107 m3, and the average annual rainfall of 1 991 mm is mainly captured during May to August (Li and Han, 2007). A small charging river has been dammed as MEX, which charges DJS occasionally in flood seasons. Water pumped from the estuary of Zhujiang River is the main water source for DJS; pumped water in 2011 varied monthly from 9.89×105 m3 to 6.47×106 m3. The maximum and minimum pumping amount occurred in September and February, respectively (Fig. 2). DJS is mesotrophic with a hydraulic retention time of 70–160 days (Table 1). FHS receives the water mainly from DJS as well as rainfall and runoff from its own small watershed. It has a similar normal level as DJS. The average annual rainfall is 1 805 mm, and the hydraulic retention time is about 60–210 days, typically longer than 100 days in most years. FHS is also mesotrophic (Table 1), while MEX is oligotrophic (Table 1). There are no charging rivers, and rainfall is the only water source. The average annual rainfall is 1 865 mm, and a small amount of flood water is charged into DJS during the wet season when the water level is higher than 21.2 m a.b.s.l (outlet elevation). In contrast to DJS and FHS, MEX is filled up solely by rainfall.

|
| Figure 2 Monthly quantity of water diversion from GCPS to DJS in 2011 |
Water samples for chemical and POM analyses were collected monthly in each lacustrine zone of the three reservoirs and one pumping station during the wet season (June 30, July 24 and August 24) and the dry season (October 26, November 24 and December 25) in 2011. Three replicates were taken near each sampling location, and the averaged value was used for analysis. Water temperature (WT), pH, dissolved oxygen (DO) and conductivity (COND) were measured using YSI sensors. Water transparency was measured using a Secchi disk. Water samples from 0.5 m below the water surface was taken, stored in an icebox and brought back to the laboratory for analysis. Phytoplankton species were counted under a microscope. Total nitrogen (TN) and total phosphorus (TP) from unfiltered water were measured spectrophotometrically. For dissolved nutrient analysis, water samples were filtered through a 0.45 μm membrane filter and determined for ammonium (NH4-+N), nitrate (NO3--N), nitrite (NO2--N) and soluble reactive phosphorus (SRP) concentrations by spectrophotometry (APHA et al., 1989). Chlorophyll a (Chl a) concentration was measured by spectrophotometry (Lorenzen, 1967; Lin et al., 2005).
Water samples for stable carbon and nitrogen isotope analysis of POM (δ13CPOM and δ15NPOM) were collected from 0.5 m below the water surface, filtered through a 100-μm plankton net to remove large zooplankton and detritus, then filtered onto precombusted (500℃ for 4 h) Whatman GF/F filters (0.7 μm) dried at 60℃ for 48 h in an oven. The filtered POM was scraped off and loaded between 0.3 and 0.5 mg into tin capsules for later analysis. Stable isotope analysis was performed using a FlashEA1112 Elemental Analyzer coupled with a Thermo Delta Plus Advantage Series Mass Spectrometer at the Nanjing Institute of Geography & Limnology, Chinese Academy of Sciences in China. The stable isotope ratios 13C/12C and 15N/14N are reported as relative to the Vienna Dee Belemnite standard (V-PDB) for δ13C and atmospheric N2 for δ15N. Analytical error was typically 0.1‰ for δ13C and 0.2‰ for δ15N.
Correlation analysis was used to demonstrate the relationship between stable isotopes of POM and environmental variables, t-test was used to compare pairwise means of chemical variables between reservoirs, and one-way analysis of variance (ANOVA) was used to compare seasonal variability in stable isotopes between reservoirs. All tests were performed using SPSS 20.0 and were considered significant at P < 0.05.
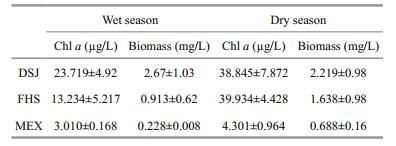
3 RESULT 3.1 Seasonal variation in physicochemical variables and phytoplanktonWater temperatures throughout the study period were above 16℃ in the three reservoirs and at the pumping station. Conductivity was highest in DJS (209 to 370 μS/cm), followed by that that in FHS (150 to 215 μS/cm) and MEX (43 to 50 μS/cm) in both wet and dry seasons. Secchi depth was similar ( < 1.0 m) in the DJS and FHS, but much higher (1.5–3 m) in MEX (Fig. 3). Both TN and DIN concentrations were not different between DJS and FHS in the wet season, and higher in the former in the dry season. For the small oligotrophic reservoir, its TN and DIN were always significantly lower than the other two well-connected reservoirs, with an increase in the dry season. TP concentration was higher in the dry season for all reservoirs, while SRP was higher in the wet season. TP was higher in DJS than in FHS and MEX in both wet and dry seasons, but not for SRP.

|
| Figure 3 Seasonal variations in select physical and chemical variables for the three reservoirs |
Phytoplankton species composition was similar between DJS and FHS in the wet and dry season, but was different from that in MEX. It was dominated by filamentous Cyanobacteria such as Pseudanabaena sp. and Planktothrix sp. in DJS and FHS and by Peridinium pusillum and Cyclotella meneghiniana in MEX. Chl a and phytoplankton biomass was higher in the dry season in all reservoirs, and was significantly higher in DJS (t=6.143, P < 0.001) and FHS (t=3.603, P=0.005) than that in MEX (Table 2).
|
MEX had the highest δ13CPOM with an average of -26.4‰, followed by the GCPS (-29.7‰). δ13CPOM was lowest in the two well-connected DJS (-33.9‰ to -29.1‰) and FHS (-30.9‰ to -33.4‰) (Fig. 4). Seasonality of δ13CPOM was similar in the two wellconnected reservoirs, i.e. significantly higher in the wet than in the dry season, but slightly different in the pumping station and in MEX. δ15NPOM had a seasonality different from δ13CPOM. Except in DJS, the δ15NPOM was higher in the wet season. MEX and FHS had lower value of δ15NPOM with a mean of 4.2‰.

|
| Figure 4 Seasonal differences in POM δ13C and δ15N ratios (mean±SD) |
When all data for wet and dry seasons are combined, δ13CPOM of DJS and MEX showed a significant difference (F=68.43, P < 0.001), and this difference also appeared between FHS, GCPS and MEX. δ15NPOM also significantly differed between DJS, FHS and MEX (F=48.979, P < 0.001), but appeared similar among FHS, MEX and GCPS (F=0.103, P=0.751).
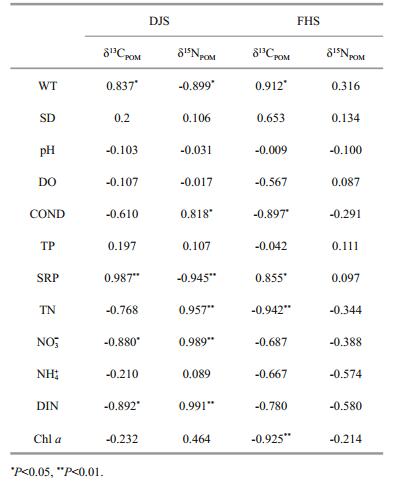
3.3 Correlation between stable isotopes of POM and environmental variablesIn DJS, δ13CPOM correlated positively with water temperature and SRP concentration, and negatively with DIN concentration. δ15NPOM correlated positively with COND, TN, NO3-, and DIN, but negatively with water temperature and SRP (Table 3). δ13CPOM in FHS correlated positively with water temperature and SRP, and negatively with COND, TN, and Chl a. No significant correlation was found between δ15NPOMand any of the environmental variables.
|
There existed a significant correlation (R2=0.256, P=0.032) for δ13CPOM between the GCPS and DJS. The δ13CPOM correlated (R2=0.750, P < 0.001) between DJS and FHS, but not between GCPS and FHS (R2=0.022, P=0.561) (Fig. 5). δ15NPOM, only significantly correlated between DJS and GCPS (R2=0.285, P=0.022) (Fig. 5). A significant relationship was found for δ15NPOM between MEX and DJS (R2=0.3, P=0.03), and no correlation between reservoirs was detected for δ13CPOM (Fig. 6).

|
| Figure 5 Similarity and dissimilarity of the seasonal variation of δ13CPOM and δ15NPOM among the well-connected reservoirs (DJS and FHS) and the GCPS |

|
| Figure 6 Similarity and dissimilarity of the seasonal variation of δ13CPOM and δ15NPOM between the well-connected reservoirs (DJS and FHS) and the relatively isolated reservoir (MEX) |
The present study demonstrates that variation of δ13CPOM and δ15NPOM in the two well-connected eutrophic reservoirs (DJS and FHS) followed seasonal hydrology. Seasonal variations of δ13CPOM and δ15NPOM are known to relate to external physical forcing factors such as temperature and photoperiod, which affect phytoplankton growth rates and isotope fractionation (Hollander and McKenzie, 1991; Lehmann et al., 2004), allochthonous loading of organic matter (Grey et al., 2001), phytoplankton species composition (Zohary et al., 1994), primary productivity (Gu et al., 2006, 2011), substrate concentrations (DIC and DIN) and their isotopic composition (Grey et al., 2000, 2001; Gu et al., 2006). The very small seasonal amplitude of POM stable isotope is typically found in lakes with low autochthonous production, and increases in eutrophic lakes in low latitude regions (Gu and Schelske, 1996; Gu et al., 2011). Temporal variation of δ13CPOM in our three reservoirs was also small throughout the study period. The amplitude of δ13CPOM ranged from 2.9‰ for MEX to 6.1‰ in FHS, and the values are correspondingly lower than those reported for subarctic and temperate lakes (Grey and Jones, 1999; Syväranta et al., 2006). The larger variation of the δ13CPOM in DJS was related to the water pumping in the dry season.
The seasonal amplitude for δ15NPOM was also small in MEX, reflecting the dominance of non-living POM in this low productivity and rain-driven reservoir. The δ15NPOM showed an inverse seasonality to the δ13CPOM in DJS, and the δ15NPOM in the dry season were higher, which is consistent with the findings of Hou et al. (2013a) in tropical reservoirs. The temporal variation of the δ15NPOM within seasons was also small in DJS but large in the FHS. The two patterns of variation for the δ15NPOM were also observed in other studies (Gu, 2009; Hadas et al., 2009).
It is well established that under high thermal energy and nutrient inputs, isotope fractionation by phytoplankton can be reduced, leading to higher δ13CPOM, that is to increasing productivity will promote δ13CPOM values (Gu et al., 2011). A recent study on a large number of low latitude reservoirs gave a similar pattern (Hou et al., 2013b). This explains why seasonal variation in δ13CPOM for both DJS and FHS closely followed variations in water temperature and SRP concentration. The small seasonal change of δ13CPOM in MEX is expected due to its low phytoplankton production and the likely dominance of non-living POM in the surface water (Gu et al., 2011). However, unlike other studies which typically found a positive relationship between Chl a and δ13CPOM, we did not find a relationship between Chl a and δ13CPOM in DJS and FHS, where high δ13CPOM was accompanied by the low Chl a in the wet season and a low δ13CPOM accompanied by a high Chl a in the dry season. This pattern may be related to both dilution effect and short retention time in our studied system, where Chl a was not accumulated despite fast growth of phytoplankton at high temperature and enriched nutrients during the wet season. By contrast, during the dry season, slowly growing phytoplankton may accumulate high biomass due to longer retention time and less dilution effect.
The lower δ13CPOM in DJS and FHS in the dry season can be due to dilution by pumping water, and this dilution may have caused a decrease in phytoplankton biomass. Primary productivity is the fundamental factor affecting aquatic inorganic C dynamics and isotope fractionation of organic matter (Gu et al., 2006). Coupled with the relatively low temperature observed in winter, the phytoplankton had a low growth rate which often coincides with isotopic depletion in POM within lakes (Hollander and McKenzie, 1991; Zohary et al., 1994; Gu et al., 2011).
The seasonally averaged δ15NPOM, being highest in DJS followed by FHS and MEX, is consistent with the findings of trophic state/productivity-mediated isotope enrichment by Gu (2009). However, the season-specific pattern of δ15NPOM is not consistent, because it was lower by an average of 2.2‰ in the wet than in the dry season in DJS (Fig. 4). A likely contributing factor to the lower δ15NPOM during the growth season is fixation of isotopically light nitrogen from the atmosphere (Lehmann et al., 2004; Gu and Schelske, 2010). As seen, the phytoplankton was dominated by filamentous Cyanobacteria such as Pseudanabaena sp. and Planktothrix sp. in DJS and FHS, which are able to fix nitrogen.
4.2 Effects of hydrological connectivity on POM isotope compositionsIn a given water, POM isotope stable composition is always regulated by autochthonous and allochthonous sources. When a lake or reservoir receives water from others, the influence of an allochthonous source is expected and depends on how much water is transported. When the allochthonous source is dominant or highly contributed, the connected waters trend to have a similar dynamic pattern (Hou et al., 2013a). In the present studied system, more water is transported from the pumping station at the river in the dry season when precipitation decreases. The effect of the river water would decrease with its indirect connectivity to the secondary receiving reservoir (FHS). The relationship between any two water bodies for δ13CPOM confirmed this prediction. A strong correlation of δ13CPOM appeared between the DJS and the pumping station, but the strongest correlation occurred between the two wellconnected reservoirs. The changing correlation is explained by different hydrodynamics in the river and reservoirs. Compared to δ13CPOM, the effect of connectivity has become weak for δ15NPOM. The correlation of δ15NPOM is negative for two wellconnected reservoirs. Nitrogen has a more complex dynamics than carbon in waters. In addition to input from the connected water, the release and resuspension of the sediments can modify the simple correlation (Pisani et al., 2016).
Compared to that in MEX, δ13CPOM in the two larger and well-connected reservoirs were depleted, suggesting there were different sources of organic matter supporting these systems. MEX is low in primary production and is driven by rainfall and terrestrial runoff. The major source of POM was likely derived from terrestrial loading. DJS also received large amounts of water from the GCPS and was affected by the low isotope composition in POM from the river water. The isotope pool of FHS is also affected more or less by the back-flowing of water from DJS. However, δ15NPOM in the FHS was considerably lower than that in the GCPS and DJS. This means some light nitrogen might be input like biological nitrogen fixation (Gu, 2009). Further study is required to account for the similarity of δ13CPOM and difference in δ15NPOM between the two well-connected reservoirs. The seasonal difference in δ15NPOM between the connected DJS and FHS may therefore be a reflection of differences in system productivity and nitrogen loading rates from agriculture runoff. δ13CPOM values in both reservoirs were affected by the supply river being depleted in 13C. By contrast, there was little seasonal fluctuation in the isotope composition of POM in MEX as it is an isolated and oligotrophic water.
5 CONCLUSIONδ13C and δ15N of POM in three tropical reservoirs show clear but different seasonal variations, which depend on how intensely the reservoirs are hydrologically connected and impacted upon by pumped water from the Zhujiang River. δ13CPOM value is low and changes slightly in the pumping station. The similar δ13CPOM values between the two wellconnected and receiving reservoirs demonstrate the influence of the pumping water. The small, less wellconnected and oligotrophic reservoir possessed a δ13CPOM that is similar to that of terrestrial C3 plants charged mainly by rainfall and terrestrial runoff. However, δ15NPOM had different dynamic patterns even in the two well-connected reservoirs, and they appear to be intricately affected by system productivity and nitrogen fixation. Both δ13CPOM and δ15NPOM in the less connected small reservoir showed little change between wet and dry seasons as might be expected for a low productive and rain-driven reservoir. Our study provides data on the isotope characteristics of tropical connected man-made waters and supports previous findings on the usefulness of stable isotope information for both limnological and hydrological analysis.
6 ACKNOWLEDGEMENTWe thank Dr. Henri Dumont from Gent University in Belgium for his reading and comments.
| American Public Health Association (APHA), American Water Works Association, Water Environment Federation. 1995. Standard methods for the examination of water and wastewater. American Public Health Association, Washington, D. C. |
| Cole J J, Carpenter S R, Kitchell J, Pace M L, Solomon C T, Weidel B, 2011. Strong evidence for terrestrial support of zooplankton in small lakes based on stable isotopes of carbon, nitrogen, and hydrogen. Proc. Natl. Acad. Sci. U.S. A., 108(5): 1975–1980. Doi: 10.1073/pnas.1012807108 |
| Grey J, Jones R I, Sleep D, 2000. Stable isotope analysis of the origins of zooplankton carbon in lakes of differing trophic state. Oecologia, 123(2): 232–240. Doi: 10.1007/s004420051010 |
| Grey J, Jones R I, Sleep D, 2001. Seasonal changes in the importance of the source of organic matter to the diet of zooplankton in Loch Ness, as indicated by stable isotope analysis. Limnol. Oceanogr., 46(3): 505–513. Doi: 10.4319/lo.2001.46.3.0505 |
| Grey J, Jones R I, 1999. Carbon stable isotopes reveal complex trophic interactions in lake plankton. Rapid Commun.Mass Spectrom., 13(13): 1311–1314. Doi: 10.1002/(ISSN)1097-0231 |
| Gu B H, Chapma A D, Schelske C L, 2006. Factors controlling seasonal variations in stable isotope composition of particulate organic matter in a soft water eutrophic lake. Limnol. Oceanogr., 51(6): 2837–2848. Doi: 10.4319/lo.2006.51.6.2837 |
| Gu B H, Schelske C L, Waters M N, 2011. Patterns and controls of seasonal variability of carbon stable isotopes of particulate organic matter in lakes. Oecologia, 165(4): 1083–1094. Doi: 10.1007/s00442-010-1888-6 |
| Gu B H, Schelske C L, 2010. Patterns and controls of nitrogen stable isotopes of particulate organic matter in subtropical lakes. Ann. Limnol. Int. J. Limnol., 46(1): 1–7. Doi: 10.1051/limn/2010005 |
| Gu B H, Schelske L, 1996. Temporal and spatial variations in phytoplankton carbon isotopes in a polymictic subtropical lake. J. Plankton Res., 18(11): 2081–2092. Doi: 10.1093/plankt/18.11.2081 |
| Gu B, 2009. Variations and controls of nitrogen stable isotopes in particulate organic matter of lakes. Oecologia, 160(3): 421–431. Doi: 10.1007/s00442-009-1323-z |
| Hadas O, Altabet M A, Agnihotri R, 2009. Seasonally varying nitrogen isotope biogeochemistry of particulate organic matter in Lake Kinneret, Israel. Limnol. Oceanogr., 54(1): 75–85. Doi: 10.4319/lo.2009.54.1.0075 |
| Han B P, Liu Z W. 2012. Tropical and Sub-tropical Reservoir Limnology in China: Theory and Practice. Springer, Netherlands. 369p. |
| Hollander D J, McKenzie J A, 1991. CO2 control on carbonisotope fractionation during aqueous photosynthesis:a paleo-pCO2 barometer. Geology, 19(9): 929–932. Doi: 10.1130/0091-7613(1991)019<0929:CCOCIF>2.3.CO;2 |
| Hou W, Gu B H, Lin Q Q, Gu J G, Han B P, 2013a. Stable isotope composition of suspended particulate organic matter in twenty reservoirs from Guangdong, southern China:implications for pelagic carbon and nitrogen cycling. Water Res., 47(11): 3610–3623. Doi: 10.1016/j.watres.2013.04.014 |
| Hou W, Gu B H, Zhang H J, Gu J G, Han B P, 2013b. The relationship between carbon and nitrogen stable isotopes of zooplankton and select environmental variables in lowlatitude reservoirs. Limnology, 14(1): 97–104. Doi: 10.1007/s10201-012-0388-2 |
| Jones R I, Grey J, Sleep D, Quarmby C, 1998. An assessment, using stable isotopes, of the importance of allochthonous organic carbon sources to the pelagic food web in Loch Ness. Proc. Roy. Soc. Lond. B, 265(1391): 105–111. Doi: 10.1098/rspb.1998.0270 |
| Kendall C, Silval S R, Kelly V J, 2001. Carbon and nitrogen isotopic compositions of particulate organic matter in four large river systems across the United States. Hydrol.Processes, 15(7): 1301–1346. Doi: 10.1002/(ISSN)1099-1085 |
| Kling G W, Fry B, O'Brien W J, 1992. Stable isotopes and planktonic trophic structure in arctic lakes. Ecology, 73(2): 561–566. Doi: 10.2307/1940762 |
| Lee J Y, Kim J K, Owen J S, Choi Y, Shin K, Jung S, Kim B, 2013. Variation in carbon and nitrogen stable isotopes in POM and zooplankton in a deep reservoir and relationship to hydrological characteristics. J. Freshwater Ecol., 28(1): 47–62. Doi: 10.1080/02705060.2012.689999 |
| Lehmann M F, Bernasconi S M, McKenzie J A, Barbieri A, Simona M, Veronesi M, 2004. Seasonal variation of the δ13C and δ15N of particulate and dissolved carbon and nitrogen in Lake Lugano:constraints on biogeochemical cycling in a eutrophic lake. Limnol. Oceanogr., 49(2): 415–429. Doi: 10.4319/lo.2004.49.2.0415 |
| Li Q H, Han B P, 2007. Structure and dynamics of phytoplankton community based CCA analysis in a pumped storage reservoir. Acta Ecol. Sinica, 27(6): 2355–2364. |
| Lin S J, He L J, Huang P S, Han B P, 2005. Comparison and improvement on the extraction method for chlorophyll a in phytoplankton. Ecol. Sci., 24(1): 9–11. |
| Lorenzen C J, 1967. Determination of chlorophyll and pheopigments:spectrophotometric equations. Limnol.Oceanogr., 12(2): 343–346. Doi: 10.4319/lo.1967.12.2.0343 |
| Pace M L, Cole J J, Carpenter S R, Kitchell J F, Hodgson J R, Van de Bogert M C, Bade D L, Kritzberg E S, Bastviken D, 2004. Whole-lake carbon-13 additions reveal terrestrial support of aquatic food webs. Nature, 427(6971): 240–243. Doi: 10.1038/nature02227 |
| Pisani O, Dodds W K, Jaffé R, 2016. Characterizing organic matter inputs to sediments of small, intermittent, prairie streams:a molecular marker and stable isotope approach. Aquat. Sci., 78(2): 343–354. Doi: 10.1007/s00027-015-0435-2 |
| Rožič P Ž, Dolenec T, Lojen S, Kniewald G, Dolenec M, 2015. Use of stable isotope composition variability of particulate organic matter to assess the anthropogenic organic matter in coastal environment (Istra Peninsula, Northern Adriatic). Environ. Earth Sci., 73(7): 3109–3118. Doi: 10.1007/s12665-014-3606-x |
| Shotbolt L A, Thomas A D, Hutchinson S M, 2005. The use of reservoir sediments as environmental archives of catchment inputs and atmospheric pollution. Prog. Phys.Geogr., 29(3): 337–361. Doi: 10.1191/0309133305pp452ra |
| Syväranta J, Hämäläinen H, Jones R I, 2006. Within-lake variability in carbon and nitrogen stable isotope signatures. Freshwater Biol., 51(6): 1090–1102. Doi: 10.1111/fwb.2006.51.issue-6 |
| Wen M, Chen C M, 2005. Characteristics and estimation of water resources variation in resent 50-years in Zhuhai City. Acta Scientiarum Naturalium Universitatis Sunyatseni, 44(S2): 272–275. |
| Yoshioka T, Wada E, Hayashi H, 1994. A stable isotope study on seasonal food web dynamics in a eutrophic lake. Ecology, 75(3): 835–846. Doi: 10.2307/1941739 |
| Zhang M, Lin Q Q, Xiao L J, Wang S, Qian X, Han B P, 2013. Effect of intensive epilimnetic withdrawal on phytoplankton community in a (sub) tropical deep reservoir. J. Limnol., 72(3). |
| Zohary T, Erez J, Gophen M, Berman-Frank I, Stiller M, 1994. Seasonality of stable carbon isotopes within the pelagic food web of Lake Kinneret. Limnol. Oceanogr., 39(5): 1030–1043. Doi: 10.4319/lo.1994.39.5.1030 |