 2017, Vol. 35
2017, Vol. 35Institute of Oceanology, Chinese Academy of Sciences
Article Information
- XU Zhengchao(许郑超), LI Caijuan(李彩娟), LING Qufei(凌去非), GAUGHAN Sarah, WANG Guocheng(王国成), HAN Xiaofei(韩晓飞)
- Early development and the point of no return in pikeperch (Sander lucioperca L.) larvae
- Chinese Journal of Oceanology and Limnology, 35(6): 1493-1500
- http://dx.doi.org/10.1007/s00343-017-6042-4
Article History
- Received Feb. 25, 2016
- accepted in principle May. 10, 2016
- accepted for publication Aug. 24, 2016




2 Department of Biology & School of Interdisciplinary Informatics, University of Nebraska at Omaha, Omaha, NE 68182-0040, USA
Pikeperch (Sander lucioperca L.) is an economically important Percidae species and is distributed throughout the basins of the Aral Sea, Black Sea, Caspian Sea and Baltic Sea in Europe, and the basins of the Ili River and the Rtysh Tixche River in China. This species is euryhaline, with a wide range of optimal temperature (0–30℃) and is a valuable aquaculture species and sport fish due to its tender meat (Schulz et al., 2007; Wang et al., 2009). Historically, the pikeperch market mainly relied on commercial fishing efforts, which resulted in a sharp decline in wild populations (Dil, 2008). To meet the need of the pikeperch market, intensive aquaculture of pikeperch has been increased in the Netherlands, Denmark, Finland, France and China (Li and Li, 1996; Fontaine, 2009; FAO, 2011). During the last decade, studies have focused on the influence of temperature on growth and gonad development (Lappalainen et al., 2009; Hermelink et al., 2011; Hermelink et al., 2013), the effect of different hormones on spawning (Rónyai, 2007; Křišt'an et al., 2013; Falahatkar and Poursaeid, 2014), and the impact of diet on the pikeperch larval digestive system (Nyina-Wamwiza et al., 2005; Kowalska et al., 2012). The major limitation of pikeperch aquaculture has been the inadequate supply of fingerlings, which is primarily the result of a high mortality during the larval stage (Mélard et al., 1996; Ruuhijärvi and Hyvärinen, 1996). In China, pikeperch eggs are hatched in tanks. After hatching, larvae are bred in the same containers during the first two weeks, and then are transferred into ponds for further culture (Li and Li, 1996). Due to the long and irregular hatching time of pikeperch larvae and simultaneous low temperature in early spring, when plankton organisms rarely form a high density, the initial ingestion time and palatable food supply in their ponds are hard to match, which leads to the low survival rate of pikeperch larvae.
Early larvae go through three developmental stages (Yin, 1995): stage 1, the endogenous nourishment stage, stage 2, the mixed nourishment stage and stage 3 the exogenous nourishment stage. At the onset of exogenous feeding, fish larvae face death from starvation if the first feeding is delayed after the PNR, which is the threshold time point when initially starved larvae lose their capability to recover from starvation, even if they are later fed ad libitum (Blaxter and Hempel, 1963). This critical point is often used as an index to experimentally quantify 'nutritional vulnerability' or 'nutritional flexibility' (Sulkin, 1978). The principal objectives of the present study have been to evaluate the timing of yolk and oil globule absorption, the initial feeding rate and the PNR of pikeperch larvae, and to provide aquaculture guidelines on the initial feeding time for pikeperch larvae.
2 MATERIAL AND METHOD 2.1 Obtaining experimental larvaeThe larvae were provided by Suzhou Shajiabang East Lake Modern Fishery Science and Technology Development Co. Ltd. The broodstock were injected with luteinizing hormone-releasing hormone analogue (LHRHa) plus domperidone hormone (DOM) (Ningbo, Renjian Pharmaceutical Co. Ltd.), with male fish receiving half the dose of female fish. Following hormone injection, broodstock were placed in a tank (4 m×2 m×1 m) with 1 female:1 male paired matching. The parent fish spawned on a round nest composed of palm. After spawning, fertilized eggs were taken every day and embryonic development was observed under a microscope. Prior to hatching, fertilized eggs were transferred to a polyvinyl chloride tank (50 cm×35 cm×30 cm). Each set of fertilized eggs were only from a single broodstock pair.
2.2 Method of cultivationThe larvae were reared in similar polyvinyl chloride tanks. During the experiment, water temperature was maintained at (15±2)℃. The larvae were divided into two groups. One group served as a control group and another group was a starvation group. All the groups were investigated in triplicate under identical conditions. The stocking density of each group was basically the same (100 ind./L). Aeration was supplied continuously and 1/3 of the water was changed every two days. The pH was 8.0±0.5 and the ammonia nitrogen remained under 0.03 mg/L. Light conditions followed the natural diurnal cycle. The control group were fed with rotifers (Brachionus diversicornis and Brachionus quadridentatus), of which density were kept at about 30 ind./mL, three times per day. The rotifers were enriched from the culture pond by a plankton net (mesh size 0.03–0.04 mm). The starvation groups were not fed and the water was filtered through a plankton net to ensure that there was no occurrence of dietary items in the water.
2.3 Morphometric characters of the larvaeThe experimental larvae hatched on March 20, 2015, considered as 0 day after hatching. Every day, 15 fish were taken from each tank of the control group and of the starvation group. After being anesthetized with MS-222 (50 mg/L), and the main characteristics of early larval development were observed by optical microscopy (Olympus). A stereomicroscope (Ji Fei company, XTL-500) and a micro camera (Yichuang Corporation, MY310), were used to measure growth trait data, including yolk-sac long-diameter (R), yolksac short-diameter (r), oil globule diameter (d), total length (L), accurate to 0.01 mm. Yolk-sac volume (VY) and the oil globule volume (Vog) were calculated as follows:
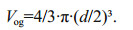
The larval oil globule and yolk-sac were combined into an ellipsoid, so yolk-sac volume was equal to the yolk-sac total volume minus the oil globule volume.
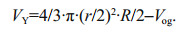
The growth rate of larvae is expressed as the specific growth rate (SGR) using the following equation: SGR=100(lnLta–lnLtb)∙t-1, where t was the time between ta and tb.
2.4 Initial feeding rate and PNREach morning at 10:00 starting from 0 DAH, 15 larvae from each tank of the starvation groups were transferred into a glass cylinder and fed with rotifers. After 2 hours, the larvae were removed, anesthetized with MS-222 (50 mg/L), then examined with a stereomicroscope to identify the presence of rotifers in their intestine. Initial feeding rate is expressed using the following equation:
Initial feeding rate=(number of larvae with rotifers in the fish intestine/ the total number of larvae)×100%.
The method to determine PNR has been adopted from Blaxter and Yin (Blaxter and Hempel, 1963; Yin and Blaxter, 1987; Yin, 1991a). After hatching, the initial feeding rate of starved larvae was determined daily. PNR was defined as the time when the initial feeding rate of the starved larvae dropped to half of the highest initial feeding rate. The experiment terminated when no starved larvae could initiate feeding.
2.5 Statistical analysisAll statistical analysis was conducted with the Statistical Product and Service Solutions 17.0. The data followed a normal distribution (Shapiro-Wilk test) to ensure that assumptions used by t-test were satisfied. Larval growth and SGR were tested by independent-samples t-test. The results are expressed as mean±SD (standard deviation) of the data, and P value less than 0.05 is considered statistically significant.
3 RESULT 3.1 Developmental characteristics of larvaeAfter hatching, the pikeperch larvae developed rapidly in morphology and experienced significant changes in their digestive system. The main morphological development characteristics were as follows.
0 DAH: the larva was straight and the whole body was transparent; the oil globule was in front of the light yellow yolk-sac (Fig. 1a).

|
| Figure 1 The growth characteristics of pikeperch (Sander lucioperca L.) larvae AC: auditory capsule; AN: anus; HG: hindgut; IN: incipient intestine; IS: intestinal secretions; IV: intestinal valve; MG: mid gut; MY: myomere; N: notochord; OG: oil globule; PF: pectoral fin; SB: swim bladder; ST: stomach; YS: yolk-sac. |
1 DAH: the larval incipient intestine was formed at the end of their yolk-sac, which was a slender fuzzy line (Fig. 1b).
2 DAH: larval sarcomere was visible and the larva possessed an open mouth (Fig. 1c).
3 DAH: the larval gut was visible. The fan-shaped pectoral fin bud was formed. The black pigment in their eyes increased, the auditory vesicle was present, and the anus was closed (Fig. 1d).
4 DAH: intestinal secretions were observed in the intestinal tract, which was followed by intestinal peristalsis (Fig. 1e).
5 DAH: the intestine began to widen and the bowel wall was smooth (Fig. 1f).
6 DAH: intestinal peristalsis had begun; the intestinal wall was slightly folded, and the anus was open with intestinal secretions being discharged (Fig. 1g).
7 DAH: the anterior part of the intestine differentiated into the early stage of the stomach.
8 DAH: the intestinal fold increased; the intestinal valve appeared; the intestine differentiated into the anterior and the posterior intestine, and intestinal peristalsis was visible. A few larvae began feeding, suggesting that the larvae went into the mixed feeding stage (Fig. 1h).
9 DAH: the bladder began to inflate, which looked slightly like the back projection, and the larvae began to swim horizontally (Fig. 1i).
10 DAH: larvae jaws moved, with enlargement of the anterior mid gut and constriction of the hindgut; black pigment fills the eyeball (Fig. 1j).
11 DAH: yolk-sac absorption was completed; the oil globule remained; the upper and lower jaws were active.
12 DAH: the digestive tract was full of food; the oil globule was oval (Fig. 1k).
14 DAH: oil globule disappeared, and the larvae began to enter the exogenous feeding stage.
16 DAH: some larvae possessed twisted spines (Fig. 1l).
18 DAH: all starved larvae had died.
3.2 The change during larval growthThe total length of newly hatched (0 DAH) pikeperch larvae was (4.25±0.22) mm. On 14 DAH, The larval total length in the control group and the starvation group was (6.05±0.14) mm and (5.92± 0.10) mm, respectively. Following the exhaustion of the yolk-sac and oil globule from 14 DAH, growth difference appeared between the control group and the starvation group (P < 0.001) (Fig. 2).

|
| Figure 2 The comparison of total length between control and starved groups of pikeperch (Sander lucioperca) larvae during 0–16 DAH Data are expressed as mean±SD and bars represent standard deviations of the means (n=15). ** indicates significant difference (P<0.01) among different groups. |
As shown in Fig. 3. From 0 to 8 DAH (endogenous feeding stage), the larval SGR in the control groups was (3.74±0.39)% and in the starvation groups (3.57±0.30)% (P>0.05). From 8 to 14 DAH (mixed feeding stage), the SGR in the control groups ((0.89±0.09)%) was significantly higher than that in the starvation groups ((0.53±0.06)%) (P < 0.05). During 14–16 DAH (exogenous feeding stage), the larvae of the starvation group demonstrated declining SGR ((2.22±0.85)%), while the larval SGR in the control groups was (3.57±0.31)%.

|
| Figure 3 The comparison of change in SGR in control and starved groups of pikeperch (Sander lucioperca) larvae during 0–16 DAH Data are expressed as mean±SD and bars represent standard deviations of the means (n=3). * indicates significant difference (P<0.05) and ** indicates significant difference (P<0.01) among different groups. |
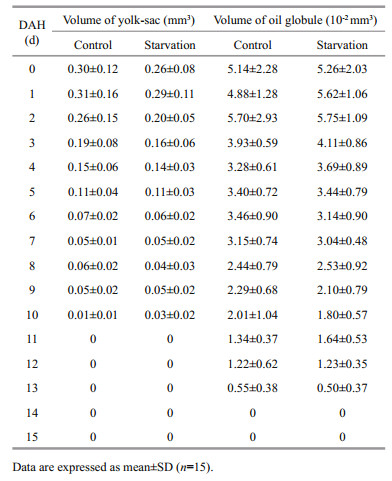
The newly hatched pikeperch larvae had a large oval yolk-sac (VY=(0.30±0.12) mm3) and a sphericalshaped oil globule (Vog=(5.14±2.28) 10-2 mm3). There was no significant difference (P>0.05) in the VY and Vog between the control and the starvation groups from 0 to 14 DAH. Complete depletion of the yolk-sac and the oil globule were respectively noticed at 11 DAH and 14 DAH, in both the control and the starved groups (Table 1).
|
Pikeperch larvae began feeding at 8 DAH, and the initial feeding rate was approximately 10%. Over the next 6 days, the initial feeding rate increased rapidly, peaking at 82.7% at 14 DAH. By 15 DAH, the initial feeding rate decreased to 48.9% and 35.6% in 16 DAH (Fig. 4). Between 15 DAH to 16 DAH, the initial feeding rate dropped to about half of the highest initial feeding rate (82.7%), and this time point should be considered as the Point of No Return.

|
| Figure 4 Initial feeding rate of pikeperch (Sander lucioperca) larvae Data are expressed as mean±SD and bars represent standard deviations of the means (n=3). |
Farris suggested that larvae can be divided into three stages: the rapid growth stage immediately following hatching, the slow growth stage around yolk-sac exhaustion and the negative growth stage following a failure to establish exogenous feeding (Farris, 1959). Based on our study, early growth of pikeperch larvae can be divided into three such stages: stage 1 (the endogenous feeding stage) is from 0 DAH to 8 DAH when the larvae were initially fed. During this stage, larval development is manifested mainly in the feeding organs, such as mouth and intestine. Larvae depend largely on yolk sources before feeding ability and physiological mechanism become fully developed. Stage 2 (the mixed feeding stage) is from 8 DAH to 14 DAH when the oil globule was exhausted. During this stage, the larval bladder begins to inflate, upper and lower jaws are active. Larval swimming and feeding ability are further improved.The beginning of the exogenous feeding facilitates the establishment of the larval movement patterns, especially cruising pattern (Yin and Blaxter, 1989). Stage 3 (the exogenous feeding stage) begins from the exhaustion of the oil globule (after 14 DAH). In stage 1, we found no significant difference in larval SGR between the control and the starvation group. After first feeding, SGR of the feeding larvae was significantly higher than that of the starved larvae. After depletion of endogenous nutrients, negative growth appeared in the starved larvae. Our results show that starvation hindered pikeperch larval growth after the endogenous feeding stage. This change is an adaptation phenomenon to improve the feeding rate and survival rate (Yin and Blaxter, 1986; Yin, 1991b).
4.2 Changes the yolk-sac and the oil globuleThe VY of newly hatched pikeperch larvae was (0.30±0.12) mm3 similar to Pomoxis nigromaculatus ((0.34±0.05) mm3) (Qin, 2014), Dicentrarchus labrax (0.30 mm3) (Rønnestad et al., 1998), and much larger than many other species, such as Lates calcarifer (0.107 mm3) (Kailasam et al., 2007) and Clupea harengus (0.182 mm3) (Yin, 1991b). The endogenous feeding of pikeperch larvae utilizes the yolk-sac and the oil globule. We found no significant differences in the depletion of either the yolk-sac or the oil globule, between the control and the starvation groups. At (15±2)℃, the pikeperch larval mixed feeding period lasted for 6 days (8–14 DAH), much longer than that of many other species such as Siniperca scherzeri (3 days, 23℃) (Zhang et al., 2009), Trachinotus ovatus (1 day, (28±1)℃) (Ma et al., 2014), and Paralichthys olivaceus (0.4 day, 14.6℃) (Dou et al., 2005). Large yolk-sacs and longer mixed feeding stages can support more nutrition reserves, which allow more time to initiate feeding before the onset of irreversible starvation (Blaxter and Hempel, 1963; Qiu et al., 2014). Of course, the time of complete yolk-sac absorption is temperature-dependent and speciesspecific (Dou et al., 2005) and to a large extent it depends on temperature (Snyder, 1976). Future studies should focus on the relation between temperature and the depletion of the yolk-sac and the oil globule.
During the mixed feeding stage, we found the yolk-sac was exhausted on 11 DAH and the oil globule became the main source in the later part of mixed feeding. This suggests that fats are utilized by pikeperch larvae less effectively and that the yolk-sac is the first and the most important source of energy (Palińska-Żarska et al., 2014), similar to in Lota lota larvae (Palińska-Żarska et al., 2014) and Lutjanus campechanus larvae (Williams et al., 2004). The presence of oil globules in some species, which could be retained for a period of time after yolk-sac exhaustion, may provide extra energy reserves and withstand starvation (Mookerji and Rao, 1999). In some teleost fishes, most of the lipids are confined to the oil globule. The size of the oil globule remains constant during embryogenesis but is reduced following yolk depletion (Silversand et al., 1996). There are two major roles for the oil globule in pikeperch larvae. Energy derived from lipid catabolism is used as energy for swimming (Rønnestad et al., 1998). The oil globule also increases larval buoyancy and allows the larvae to achieve a vertical position in the water to facilitate feeding (Hunt et al., 1996; Williams et al., 2004; Palińska-Żarska et al., 2014).
4.3 The initial feeding rate and PNRAccording to the duration of highest initial feeding rate, larvae can be divided into two types. Type A larvae start to feed at a low initial feeding rate followed by a rapid increase and then a successively rapid decrease, similar to Pagrosomus major (Bao et al., 1998) and Acipenser sinensis (Chai et al., 2011). Type B larvae possess a low initial feeding rate which rapidly increases and is maintained for a period of time, similar to Paralichthys olivaceus (Dou et al., 2005) and Gadus morhua (Overton et al., 2010). The duration of the highest feeding rate is proportional to the larval resistance to starvation. Pikeperch larvae can be classified as a Type A. Our results show that pikeperch larvae reached a maximum initial feeding rate at 14 DAH which decreased rapidly within 24 hours. Moreover, oil globule exhaustion was completed on 14 DAH, and shortly thereafter larvae entered the PNR (15–16 DAH). If there is no food, the pikeperch larvae can withstand starvation for only 1–2 days at (15±2)℃. Paralichthys olivaceus larvae took 2–3.7 days from yolk exhaustion to PNR at 15– 21℃ (Dou et al., 2005); Clupea harengus larvae took 3–5 days from yolk exhaustion to PNR at 7.5–13.1℃ (Blaxter and Hempel, 1963; Yin and Blaxter, 1987); Miichthys miiuy larvae took 2 days from yolk exhaustion to PNR at 24℃ (Shan et al., 2009). The ability of pikeperch larvae to withstand starvation was weaker than other species and highlights the need to identify the critical initial feeding time to maximize aquaculture efforts.
5 CONCLUSIONThis study has shown that at (15±2)℃, the appropriate first feeding time for pikeperch larvae is 11–12 DAH, when the initial feeding rate is higher than half of the maximum initial feeding rate. If the first feeding is later than 15 DAH, the pikeperch larvae have entered the PNR, which will result in mortality.
| Bao B L, Su J X, Yin M C, 1998. Effect of delayed feeding on feeding ability, survival and growth of red sea bream and olive flounder larvae during early development. J. Fish. China, 22(1): 33–38. |
| Blaxter J H S, Hempel G, 1963. The influence of egg size on herring larvae (Clupea harengus L.).. ICES J. Mar. Sci., 28(2): 211–240. Doi: 10.1093/icesjms/28.2.211 |
| Chai Y, Xie C, Wei Q W, 2011. Yolk-sac absorption and point of no return in Chinese sturgeon Acipenser sinensis larvae. J. Appl. Ichthyol., 27(2): 687–689. Doi: 10.1111/j.1439-0426.2010.01631.x |
| Dil H. 2008. The European market of the pikeperch for human consumption. In: Fontaine P, Kestemont P, Teletchea F, Wang N eds. Proceedings of Percid Fish Culture from Research to Production. Presses Universitaires de Namur, Namur, Belgium. p. 15-16. http://akademiai.com/doi/abs/10.1556/ABiol.58.2007.2.6 |
| Dou S Z, Masuda R, Tanaka M, Tsukamoto K, 2005. Effects of temperature and delayed initial feeding on the survival and growth of Japanese flounder larvae. J. Fish Biol., 66(2): 362–377. Doi: 10.1111/jfb.2005.66.issue-2 |
| Falahatkar B, Poursaeid S, 2014. Effects of hormonal manipulation on stress responses in male and female broodstocks of pikeperch Sander lucioperca. Aquacult.Int., 22(1): 235–244. Doi: 10.1007/s10499-013-9678-x |
| FAO. 2011. Cultured aquatic species information programme Sander lucioperca (Linnaeus, 1758). http://www.fao.org/fishery/culturedspecies/Sander_lucioperca/en . |
| Farris D A, 1959. A change in the early growth rates of four larval marine fishes. Limnol. Oceanogr., 4(1): 29–36. Doi: 10.4319/lo.1959.4.1.0029 |
| Fontaine P, 2009. Development of European inland fish culture and domestication of new species. Cah. Agric., 18(2): 144–147. |
| Hermelink B, Wuertz S, Rennert B, Kloas W, Schulz C, 2013. Schulz C. 2013.Temperature control of pikeperch (sander lucioperca)maturation in recirculating aquaculture systems-inductionof puberty and course of gametogenesis. Aquaculture, 400-401: 36–45. Doi: 10.1016/j.aquaculture.2013.02.026 |
| Hermelink B, Wuertz S, Trubiroha A, Rennert B, Kloas W, Schulz C, 2011. Influence of temperature on puberty and maturation of pikeperch. Sander lucioperca. Gen. Comp.Endocrinol., 172(2): 282–292. Doi: 10.1016/j.ygcen.2011.03.013 |
| Hunt I V H, Boutilier R G, Miyake T, Hall B K, 1996. Effects of temperature on morphological landmarks critical to growth and survival in larval Atlantic cod (Gadus morhua). Mar. Biol., 124(4): 593–606. Doi: 10.1007/BF00351041 |
| Kailasam M, Thirunavukkarasu A R, Selvaraj S, Stalin P, 2007. Effect of delayed initial feeding on growth and survival of Asian sea bass Lates calcarifer (Bloch) larvae. Aquaculture, 271(1-4): 298–306. Doi: 10.1016/j.aquaculture.2007.05.005 |
| Kowalska A, Zakęś Z, Siwicki A K, Jankowska B, Jarmołowicz S, Demska-Zakęś K, 2012. Impact of diets with different proportions of linseed and sunflower oils on the growth, liver histology, immunological and chemical blood parameters, and proximate composition of pikeperch Sander lucioperca (L).. Fish Physiol. Biochem., 38(2): 375–388. Doi: 10.1007/s10695-011-9514-z |
| Křišt'an J, Alavi S M H, Stejskal V, Policar T, 2013. Hormonal induction of ovulation in pikeperch (Sander lucioperca L.) using human chorionic gonadotropin (hCG) and mammalian GnRH analogue.. Aquacult. Int., 21(4): 811–818. Doi: 10.1007/s10499-012-9572-y |
| Lappalainen J, Milardi M, Nyberg K, Venäläinen A, 2009. Effects of water temperature on year-class strengths and growth patterns of pikeperch (Sander lucioperca (L.)) in the brackish Baltic Sea.. Aquat. Ecol., 43(1): 181–191. Doi: 10.1007/s10452-007-9150-y |
| Li R Q, Li Z R, 1996. The biological habit and breeding situation of pikeperch (Sander lucioperca L.).. J. Beijing Fisheries(S2): 23–24. |
| Ma Z H, Guo H Y, Zheng P L, Wang L, Jiang S G, Qin J G, Zhang D C, 2014. Ontogenetic development of digestive functionality in golden pompano Trachinotus ovatus(Linnaeus 1758). Fish Physiol. Biochem., 40(4): 1157–1167. |
| Mélard C, Kestemont P, Grignard J C, 1996. Intensive culture of juvenile and adult Eurasian perch (P.fluviatilis):effect of major biotic and abiotic factors on growth.. J. Appl.Ichthyol., 12(3-4): 175–180. Doi: 10.1111/jai.1996.12.issue-3-4 |
| Mookerji N, Rao T R, 1999. Rates of yolk utilization and effects of delayed initial feeding in the larvae of the freshwater fishes rohu and singhi. Aquacult. Int., 7(1): 45–56. Doi: 10.1023/A:1009244819835 |
| Nyina-Wamwiza L, Xu X L, Blanchard G, Kestemont P, 2005. Effect of dietary protein, lipid and carbohydrate ratio on growth, feed efficiency and body composition of pikeperch sander lucioperca fingerlings. Aquacult. Res., 36(5): 486–492. Doi: 10.1111/are.2005.36.issue-5 |
| Overton J L, Meyer S, Støttrup J G, Peck M A, 2010. Role of heterotrophic protists in first feeding by cod (Gadus morhua) larvae. Mar. Ecol. Prog. Ser., 410: 197–204. Doi: 10.3354/meps08658 |
| Palińska-Żarska K, Żarski D, Krejszeff S, Nowosad J, Biłas M, Trejchel K, Kucharczyk D, 2014. Dynamics of yolk sac and oil droplet utilization and behavioural aspects of swim bladder inflation in burbot, Lota lota, L, larvae during the first days of life, under laboratory conditions.. Aquacult. Int., 22(1): 13–27. Doi: 10.1007/s10499-013-9663-4 |
| Qin Z Q, 2014. Experimental starvation on Pomoxis nigromacufatus larvae and definition of the point of no return. Fujian J. Agric. Sci., 29(12): 1167–1173. |
| Qiu C G, Xu S L, Lin S Z, Qi C, 2014. Effects of delayed feeding on survival and growth of Sebastiscus marmoratus larva. J. Ningbo Univ. (NSEE), 27(3): 7–11. |
| Rønnestad I, Koven W, Tandler A, Harel M, Fyhn H J, 1998. Utilisation of yolk fuels in developing eggs and larvae of European sea bass (Dicentrarchus labrax). Aquaculture, 162(1-2): 157–170. Doi: 10.1016/S0044-8486(98)00203-8 |
| Rónyai A, 2007. Induced out-of-season and seasonal tank spawning and stripping of pike perch (Sander lucioperca L).. Aquacult. Res., 38(11): 1144–1151. Doi: 10.1111/are.2007.38.issue-11 |
| Ruuhijärvi J, Hyvärinen P, 1996. The status of pike-perch culture in Finland. J. Appl. Ichthyol., 12(3-4): 185–188. Doi: 10.1111/jai.1996.12.issue-3-4 |
| Schulz C, Böhm M, Wirth M, Rennert B, 2007. Effect of dietary protein on growth, feed conversion, body composition and survival of pike perch fingerlings(Sander lucioperca). Aquacult. Nutr., 13(5): 373–380. Doi: 10.1111/anu.2007.13.issue-5 |
| Shan X J, Cao L, Huang W, Dou S Z, 2009. Feeding, morphological changes and allometric growth during starvation in miiuy croaker larvae. Environ. Biol. Fish., 86(1): 121–130. Doi: 10.1007/s10641-008-9412-0 |
| Silversand C, Norberg B, Haux C, 1996. Fatty-acid composition of ovulated eggs from wild and cultured turbot(Scophthalmus maximus) in relation to yolk and oil globule lipids. Mar. Biol., 125(2): 269–278. Doi: 10.1007/BF00346307 |
| Snyder D E. 1976. Terminologies for intervals of larval fish development. In: Great Lakes Fish Egg and Larvae Identification: Proceedings of a Workshop. U. S. Fish Wildl. Serv., Michigan. p. 41-60. |
| Sulkin S D, 1978. Nutritional requirements during larval development of the portunid crab, Callinectes sapidus Rathbun. J. Exp. Mar. Biol. Ecol., 34(1): 29–41. Doi: 10.1016/0022-0981(78)90055-2 |
| Wang N, Xu X L, Kestemont P, 2009. Effect of temperature and feeding frequency on growth performances, feed efficiency and body composition of pikeperch juveniles(Sander lucioperca). Aquaculture, 289(1-2): 70–73. Doi: 10.1016/j.aquaculture.2009.01.002 |
| Williams K, Papanikos N, Phelps R P, Shardo J D, 2004. Development, growth, and yolk utilization of hatcheryreared red snapper lutjanus campechanus larvae. Mar.Ecol. Prog. Ser., 275: 231–239. Doi: 10.3354/meps275231 |
| Yin M C, Blaxter J H S, 1986. Morphological changes during growth and starvation of larval cod (Gadus morhua L) and flounder (Platichthysflesus L.). . J. Exp. Mar. Biol.Ecol., 104(1-3): 215–228. Doi: 10.1016/0022-0981(86)90106-1 |
| Yin M C, Blaxter J H S, 1987. Feeding ability and survival during starvation of marine fish larvae reared in the laboratory. J. Exp. Mar. Biol. Ecol., 105(1): 73–83. Doi: 10.1016/S0022-0981(87)80030-8 |
| Yin M C, Blaxter J H S, 1989. Cruising speeds during early development and starvation of marine fish larvae. Oceanol.Limnol. Sin., 20(1): 1–9. |
| Yin M C, 1991a. Advances and studies on early life history of fish. J. Fish. China, 15(4): 348–358. |
| Yin M C, 1991b. Feeding ability and growth of the yolk-sac larvae of North Sea herring. Oceanol. Limnol. Sin., 22(6): 554–560. |
| Yin M C, 1995. Feeding and growth of the larva stage of fish. J. Fish. China, 19(4): 335–342. |
| Zhang L, Wang Y J, Hu M H, Fan Q X, Chenung S G, Shin P K S, Li H, Cao L, 2009. Effects of the timing of initial feeding on growth and survival of spotted mandarin fish Siniperca scherzeri larvae. J. Fish Biol., 75(6): 1158–1172. Doi: 10.1111/jfb.2009.75.issue-6 |