 2018, Vol. 36
2018, Vol. 36Institute of Oceanology, Chinese Academy of Sciences
Article Information
- CHEN Yunyan(陈芸燕), SUN Xiaoxia(孙晓霞), ZHUN Mingliang(朱明亮)
- Net-phytoplankton communities in the Western Boundary Currents and their environmental correlations
- Chinese Journal of Oceanology and Limnology, 36(2): 305-316
- http://dx.doi.org/10.1007/s00343-017-6261-8
Article History
- Received Sep. 29, 2016
- accepted in principle Dec. 20, 2016
- accepted for publication Dec. 26, 2016




2 University of Chinese Academy of Sciences, Beijing 100049, China;
3 Laboratory of Marine Ecology and Environmental Science, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China
Western Boundary Currents, which are the most intensive transport features in the world, advect warm nutrient poor waters into more polar areas and displace productive waters (Loder et al., 1998; Koslow et al., 2008; Everett et al., 2014). The Western Boundary Currents are often thought to be unproductive compared to the Eastern Boundary Currents (Everett et al., 2014). The oceanography features of the Western Boundary Currents have been extensively studied (Lukas et al., 1991; Hu et al., 2015) and their circulations are illustrated in Fig. 1. In Western Boundary Currents, the link between oceanographic processes and phytoplankton communities is complex due to the combination of wind and current-driving upwellings (Everett et al., 2014). It has also been reported that a large proportion of the tropical and subtropical ocean is well stratified for most of the year, which restricts nutrient supply from deeper water and turns the surface environment oligotrophic (Shiozaki et al., 2013). Phytoplankton blooms can occasionally occur as a result of upwellings associated with current meandering (Christian et al., 2004), but biological production shows little seasonal variation because of the oligotrophic conditions that persist throughout the year (Le Borgne et al., 2002).

|
| Figure 1 Study area and locations of the sampling stations in the north-western Pacific during the cruise from 23 August to 29 September, 2014 Map of the north-western Pacific shows the major geographic names and the surface currents, including the Kuroshio Current (KC), the Luzon Current (LC), the Mindanao Current (MC), the North Equatorial Current (NEC), the North Equatorial Counter Current (NECC), the Halmahera Eddy (HE), and the Mindanao Eddy (ME) (Lukas et al., 1991; Hu et al., 2015). |
Marine plankton are usually strongly affected by mesoscale hydrographic features, such as fronts and eddies (Rodríguez et al., 1999; Okazaki et al., 2002). Cleve (1897) and Aurivillius (1898) proposed that some species of plankton were indigenous to specific water masses and could be used to indicate water movements because many marine species are transported by oceanic currents (Thorson, 1950; Moita et al., 2016). Physical transport and dispersal has been recognized as a population-structuring mechanism in ecosystems (Shmida and Wilson, 1985; Chesson, 2000). For this reason, the distribution of many phytoplankton species corresponds closely to the restrictive environments found in many marine ecosystems. Phytoplankton communities play a vital role in biogeochemical cycles and the pelagic food webs, and they also support marine ecosystem energy demands (Wang et al., 2016). They are widely distributed in the epipelagic layer and the distribution of phytoplankton species and their community structures are usually associated with the major oceanic environments (Ahmed et al., 2016; Carney et al., 2016). In phytoplankton communities, the diatom species, which have a specific sensitivity to ecological characteristics (Stevenson et al., 1999; Gomà et al., 2005; Salomoni et al., 2006), are often used as bioindicators (Lacuna, 2003). In general, the epipelagic phytoplankton communities are usually affected by temperature, light conditions, nutrients, and predation by zooplankton and fish (Tilman et al., 1982; Jiang et al., 2014).
This study collected quantitative information from 69 net samples about the ecology of oceanic phytoplankton species in the Western Boundary Currents. We focused on (1) the spatial variability of the phytoplankton community (composition, abundance, and biomass) in the studied region; (2) how the physical oceanographic processes maintain and modulate phytoplankton abundance and biomass; and (3) how phytoplankton abundance spatial variability is related to environmental factors.
2 MATERIAL AND METHOD 2.1 Study area and strategyThe summer 2014 cruise on board the RV KEXUE took place in the north-western Pacific between August 23rd and September 29th, 2014. The area was delimited by the coordinates 0°–21.5°N, 121°– 135.5°E (Fig. 1). Fifty-five stations were sampled at between 0 and 300 m depth by vertical tows with a conical plankton net (mouth opening 0.1 m2, mesh size 77 μm). Phytoplankton samples were immediately preserved in 5% buffered formalin-seawater solution. The seawater was collected with a rosette sampler that carried 24 (10 dm3) Niskin bottles and CTD sensors (SBE 11 plus). At all stations, temperature and salinity vertical profiles were measured using a seabird 911 CTD, and seawater was collected at six depths (1, 30, 75, 100, 150, and 300 m) for nutrients and chlorophyll a (Chl a) analysis.
2.2 Chlorophyll aThe seawater samples (1 L) that were used to determine total Chl a concentrations were filtered through Whatman GF/F filters (0.7 μm). Chl a was subsequently extracted in 10 cm3 of 90% acetone for 24 h at 20℃. Then the Chl a concentrations were measured using a Turner fluorimeter (Model 10-AU) (Yentsch and Menzel, 1963; Lorenzen, 1966).
2.3 NutrientsNutrient measurements (NO3-N, NO2-N, and PO3-P) were performed on samples taken from every station. The samples for each depth level were frozen at -20℃ for further analysis. This was carried out according to Lu et al. (2015) using a nutrient continuous flow analyzer (SKALAR San++, Breda, the Netherlands). The detection limits were 50, 50, and 20 nmol/L for NO3-N, NO2-N, and PO4-P, respectively.
2.4 Phytoplankton identification and quantificationFor microscopy, 69 phytoplankton net samples were stored in the lab for 2 months. The species composition and abundances of the phytoplankton were measured using an optical digital biological microscope (Olympus 13X51) at 100× magnification in the laboratory. The phytoplankton species were identified following Omura et al. (2012).
2.5 Carbon biomass calculationThe method described by Yang et al. (2016) was found to be suitable for the phytoplankton net samples and the relationships were as follows:
for diatoms:
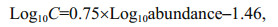 (1)
(1)for dinoflagellates:
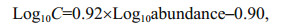 (2)
(2)where C represents the amount of carbon (pg).
Trichodesmium carbon biomass was computed using 0.011 μg C/trichome as the conversion factor (Carpenter, 1983).
2.6 Statistical analysisCluster analysis. Cluster analysis was used to examine the spatial variation in the phytoplankton species composition. Species abundance data were fourth-root transformed prior to analysis. The BrayCurtis (1957) similarity index was used to compare species composition between samples. The similarity indices were coupled using hierarchical agglomerative clustering and the group average method was applied. Similarity percentage analysis was performed to compare the variation in species composition among communities. All these analyses were carried out using PRIMER version 5 for Windows (PRIMER-E Ltd., Plymouth, UK).
Relationship between phytoplankton community structure and environmental variables. Principal Component Analysis (PCA) was used to visualize the relationships between the abundance of different phytoplankton taxa and environmental factors (temperature, salinity, nitrate+nitrite, phosphorus and Chl a concentrations) in the different sampling areas. The correlation coefficients were calculated to identify any relationships between phytoplankton abundance and the physicochemical factors. The Kaiser-Meyer-Olkin's (KMO) measure of sampling adequacy and Bartlett's test of sphericity were used to assess the usefulness and adequacy of the PCA (Budaev, 2010). All these statistical analyses were performed using STAT (Version 11 for Windows, STAT, USA). Pearson correlation analysis was also used to test the significance of the relationships between phytoplankton (abundance and biomass) and the environmental factors.
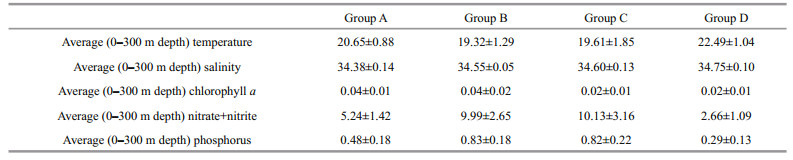
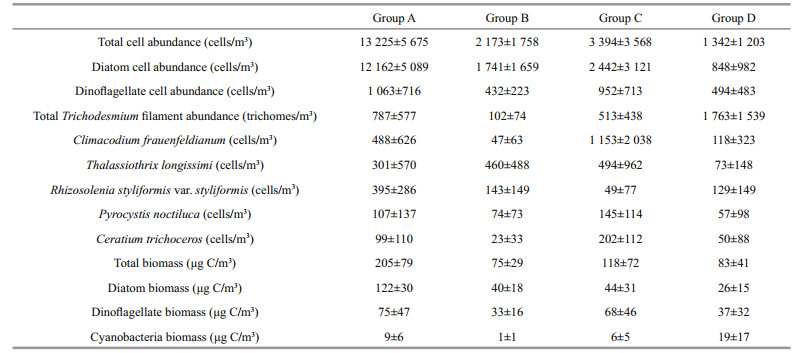
3 RESULT 3.1 Hydrology, nutrients, and chlorophyll analyses of the Western Boundary CurrentsFive transects (T1, T2, T3, T4, and T5) in the Western boundary Currents were surveyed in this study (Fig. 1). The cluster analysis identified four phytoplankton groups (Fig. 2). The physical properties of the water column were used to investigate how the different groups were influenced by different currents. These properties were temperature (T), salinity (S), nitrate+nitrite (N), phosphorus (P), and Chl a. The water samples for Group D were warm, saline, and oligotrophic, (xT=22.49±1.04℃, xS=34.75±0.10, xN=2.66±1.09 μmol/L, xP=0.29±0.13 μmol/L, and xChl a=0.02±0.01 mg/m3) (Fig. 3, Table 1); the lowest average temperature and high average nutrients occurred in the samples for Group B with xT=19.32±1.29℃, xN=9.99±2.65 μmol/L, and xP=0.83±0.18 μmol/L (Fig. 3, Table 1); and the highest average nutrients occurred in the water samples for Group C with xN=10.13±3.16 μmol/L and xP=0.82±0.22 μmol/L (Fig. 3, Table 1). Group A water samples had the lowest salinity and the highest Chl a, with xS=34.38±0.14 and xChl a=0.04±0.01 mg/m3 (Fig. 3, Table 1).

|
| Figure 2 Location of the different phytoplankton groups in the north-western Pacific The four phytoplankton groups: Group A, Group B, Group C, and Group D, were divided according to cluster analysis performed on the phytoplankton abundance data sets. |

|
| Figure 3 Temperature and salinity (T-S) relationships at 0–300 m depth in the four phytoplankton groups (Group A, Group B, Group C, and Group D) |
|
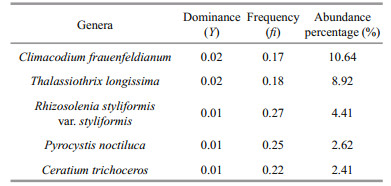
The phytoplankton net samples obtained from the north-western Pacific were analyzed and 199 species belonging to 61 genera in four phyla were identified. There were 114 diatoms belonging to 45 genera (57.29% of total species), 82 species in 14 genera of dinoflagellate (41.21%), two cyanobacteria species, and one Chrysophyta species (Table 2). The phytoplankton in the survey region mostly consisted of single celled or filamentous organisms. The filamentous phytoplankton were mainly composed of Trichodesmium spp., but the filamentous phytoplankton abundance, which was calculated as the number of trichomes/m3, was not included in the total phytoplankton cell abundance count (cells/m3). The diatom abundance accounted for 64.66% of the total phytoplankton cell abundance, whereas dinoflagellates accounted for 35.31% (Fig. 4a). The two cyanobacteria species were distributed widely across the survey region (Fig. 4b). The phytoplankton species classified as dominant in the survey region were mainly eurythermic diatoms, but sometimes highly saline tropical dinoflagellates and oceanic cyanobacteria were also classified as dominant. They included Climacodium frauenfeldianum (10.64%), Thalassiothrix longissima (8.92%), Rhizosolenia styliformis var.styliformis (4.41%), Pyrocystis noctiluca (2.62%), and Ceratium trichoceros (2.41%) (Table 3). Trichodesmium thiebautii accounted for 99.11% of the cyanobacteria found (Fig. 4b). The diatom C. frauenfeldianum is an oceanic species commonly found in tropical or subtropical seas (Carpenter and Janson, 2000); T. longissimi is an oceanic diatom and has a worldwide distribution (Hasle and Semina, 1987; Reid et al., 1992); R. styliformis var. styliformis is an oceanic diatom with a worldwide distribution (Brown and Werner, 1978), P. noctiluca is a large, autotrophic, relatively slowgrowing vacuolar dinoflagellate that is abundant in tropical and subtropical ocean netplankton (Taylor, 1973; Rivkin et al., 1982); and C. trichoceros is an autotrophic dinoflagellate that is abundant in tropical ocean netplankton (Hallegraeff and Jeffrey, 1984).

|

|
| Figure 4 Horizontal distribution of phytoplankton abundance and carbon biomass in the Western Boundary Currents a. phytoplankton cell abundance (cells/m3); b. Trichodesmium spp. filament abundance (trichomes/m3); c. phytoplankton carbon biomass (μg C/m3); and d. dominant species abundance (cells/m3). |
The cluster analysis results were used to analyze similarities among phytoplankton taxa based on their abundance during the 2014 summer cruise. Four distinct phytoplankton communities were revealed by the cluster analyses (Fig. 2). Phytoplankton groups that contained fewer than four stations were not considered significant clusters. Clusters were assigned symbols with different colors and plotted at the sampling region (Fig. 2). The cluster analyses showed that the phytoplankton community had a spatial distribution structure in the Western Boundary Currents.
Group A contained stations distributed in the Luzon Strait (Fig. 2), which is the main water exchange channel between the western Pacific and the South China Sea (Tang et al., 1999). It had the highest phytoplankton abundance (13 225 cells/m3) and biomass (205 μgC/m3) among the four communities (Fig. 4a, c; Table 4). The diatoms contributed 59% to the total biomass, the dinoflagellates contributed 37%, and the cyanobacteria contributed 4% (Table 4). The dominant species, R. styliformis var. styliformis, had the highest phytoplankton abundance (395 cells/m3) in Group A (Fig. 4d; Table 4).
|
Group B contained stations that are affected by the North Equatorial Current (NEC) and the south flowing Mindanao Current (MC) (Fig. 2). Group B had the lowest Trichodesmium spp. abundance (102 trichomes/m3) and biomass (75 μgC/m3) among the four communities (Fig. 4a, c; Table 4). The diatoms contributed 54% to the total biomass, the dinoflagellates contributed 44%, and the cyanobacteria contributed 2% (Fig. 4d; Table 4).
Group C contained stations affected by the North Equatorial Counter Current (NECC) (Fig. 2), and had a relatively high phytoplankton abundance (3 394 cells/m3) and biomass (118 μg C/m3) (Fig. 4a, c; Table 4). The diatoms contributed 37% to the total biomass, the dinoflagellates contributed 58% to the total biomass, and the cyanobacteria contributed 5% to the total biomass (Table 4). The dominant species C. frauenfeldianum (1 153 cells/m3), T. longissima (494 cells/m3), P. noctiluca (145 cells/m3), C. trichoceros (202 cells/m3) had the highest phytoplankton abundances in Group C (Fig. 4d; Table 4).
Group D mainly contained the stations affected by the Subtropical Counter Current (STCC) and the northward flowing Kuroshio Current (KC) (Fig. 2). Group D had the lowest phytoplankton cell abundance (1 342 cells/m3) among the four communities (Fig. 4a, c; Table 4), whereas Trichodesmium filament abundance (1 763 trichomes/m3) was high in Group D (Table 4). The diatoms contributed 32% to the total biomass, the dinoflagellates contributed 45%, and the cyanobacteria contributed 24% (Table 4).
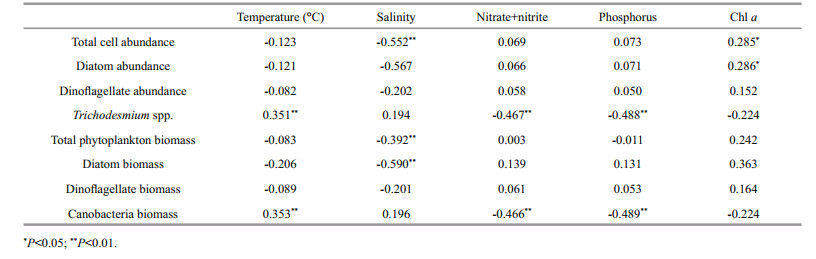
3.4 Phytoplankton distribution in relation to environmental factorsThe KMO's measure of sampling adequacy (0, 704) and Bartlett's test of sphericity (P < < 0.000 1) indicated the usefulness of the PCA. The PCA determined three major components that explained 34.9%, 21.2% and 10.8% of the total variability, respectively (Fig. 5a). The first component was significantly correlated with eleven out of the thirteen variables: T. longissima cell abundance, diatom cell abundance, P. noctiluca cell abundance, T. thiebautii filament abundance, C. frauenfeldianum cell abundance, C. trichoceros cell abundance, average (0–300 m depth) temperature, average (0–300 m depth) salinity, average (0–300 m depth) nitrate+nitrite, average (0–300 m depth) phosphorus, average (0–300 m depth), and chlorophyll a (except T. thiebautii filament abundance). All the variables were positively correlated. The second component was significantly correlated with dinoflagellate cell abundance and R. styliformis var. styliformis cell abundance, but moderately correlated with diatom cell abundance, P. noctiluca cell abundance, and C. trichoceros cell abundance. All four variables were positively correlated (Fig. 5a). The graphical representation (Fig. 5b) constructed from the first two principal components and the cluster analysis (Fig. 2) showed similar results. In summary, this analysis suggests greater T. thiebautii filament abundances in the STCC and LC regions were associated with higher water temperature and salinity conditions. The increased diatom, C. frauenfeldianum, T. longissima, P. noctiluca, and C. trichoceros cell abundances in the NECC region were associated higher nutrient conditions.

|
| Figure 5 Hierarchical clustering illustrations for the first two principal components of the Principal Component Analysis (PCA) performed on the various parameters measured These were average (0-300 m depth) temperature (T), average (0-300 m depth) salinity (S), average (0-300 m depth) nitrate+nitrite (N), average (0-300 m depth) phosphorus (P), average (0-300 m depth) Chl a, diatom cell abundance (1), dinoflagellate cell abundance (2), Trichodesmium thiebautii filament abundance (3), Climacodium frauenfeldianum cell abundance (4), Thalassiothrix longissima cell abundance (5), Rhizosolenia styliformis var.styliformis cell abundance (6), Pyrocystis noctiluca cell abundance (7), and Ceratium trichoceros cell abundance (8). a. the circle indicates the first two dimensions of the PCA; b. the analysis classified the sampling stations into four clusters: the first incorporated the LC region (Group A), the second incorporated the NEC region (Group B), the third incorporated the NECC region (Group C) and the fourth incorporated the STCC region (Group D). |
This study showed that the latitudinal distribution of Trichodesmium in the western Pacific Ocean was asymmetric with higher abundances on the subtropical side. Our results were consistent with previous observations in the tropical and subtropical Pacific Ocean (Yang et al., 2002; Kitajima et al., 2009; Jiang et al., 2015). Strong temperature and salinity fronts, which divided Group C and Group D, were observed near 16°N during the summer cruise (Nan et al., unpublished). The salinity front between 18.0°N and 15.9°N was associated with the NEC (Kimura and Tsukamoto, 2006; Kitajima et al., 2009). In addition, higher N2 fixation activity was observed between the Kuroshio Extension and the salinity front in the NEC, but the equatorial waters showed low N2 fixation activities over the summer (Kitajima et al., 2009).
Trichodesmium inhabits the surface waters of oligotrophic, tropical, and subtropical oceans, and its abundance is especially high in the Western Boundary Currents (Capone et al., 1997). However, to date, it has not been clear how environmental factors control the latitudinal distribution of Trichodesmium. Our results suggest that temperature is an important physiological restriction on the geographic distribution of Trichodesmium (Fig. 5a, Table 5), which is similar to previous observations of the tropical and subtropical oceans (Capone et al., 1997; Zehr and Ward, 2002; Breitbarth et al., 2007; Stal et al., 2010). Studies on the tropical and subtropical oceans have shown that the optimum temperature for N2 fixation and growth in the oceans lies within the range 20℃ and 30℃ (Capone et al., 1997; Breitbarth et al., 2007). Significant positive correlations between Trichodesmium abundance and water temperature were recorded in our survey (Fig. 5a, Table 5). However, Trichodesmium could be transported to higher latitudes with low temperatures by oceanic currents (Carpenter, 1983; Lipschultz and Owens, 1996). Therefore, Trichodesmium distribution was probably directly influenced by water temperature and current.
|
This survey provided evidence for the crucial role of temperature in the distribution of Trichodesmium across the western Pacific Ocean. However, Trichodesmium was less abundant in the high surface water temperature areas found in the NECC and NEC regions (Fig. 4b, Table 4). The absence of diazotrophs at the equator has often been mentioned in previous research (Stal, 2009). Hood et al. (2004) also found that the association between Trichodesmium abundance and temperature was generally attributable to oceanographic features associated with warm waters rather than a direct physiological response to temperature itself. This means that other factors are probably involved in determining the distribution of Trichodesmium in the open sea. Previous studies have suggested that nutrient supply seems to be the main mechanism for eliminating the usually high diazotroph abundance levels (Stal, 2009). The high concentration of nitrogen in warm water regions would promote other phytoplankton over diazotrophic organisms (Langlois et al., 2008; Stal, 2009). This is also suggested by the relatively high nitrogen levels in the NEC and NECC regions (Table 1). The correlation between Trichodesmium biomass and nitrogen concentration was significantly negative (Table 5). However, Trichodesmium abundance was high in the Luzon Strait station samples (Fig. 4b, Table 4) where there is effective water exchange between the western Pacific and the South China Sea (Shaw and Chao, 1994; Hu et al., 2000; Chen et al., 2003). The high Trichodesmium abundance may be caused by the Kuroshio Current that flows from the oligotrophic STCC region. Phosphorus has also been identified as a possible nutrient that limits Trichodesmium abundance (Sañudo-Wilhelmy et al., 2001; Voss et al., 2004). In this survey, the phosphorus levels in the tropical and subtropical regions were low (Table 1), which confirmed previous studies that suggested that phosphorus consumption may have been enhanced by N2 fixation (Tyrrell, 1999; Hashihama et al., 2009).
4.2 The relationships between upwellings, eddies, and phytoplankton communitiesIn tropical and subtropical ocean regions, phytoplankton abundance and biomass are influenced by current-driven upwelling events and eddies (Vaillancourt et al., 2003; Pennington et al., 2006; Huang et al., 2010; Bibby and Moore, 2011). The results from the NECC region stations showed that there was a high phytoplankton abundance and biomass in our study area (Table 4). There is a considerable upwelling of low temperature and low salinity waters from 5°N to 10°N (Nan et al., unpublished). Our results suggested that the phytoplankton bloom along the NECC was mostly caused by the upwelling associated with current meandering, which was consistent with previous observations in the equatorial Pacific (Christian et al., 2004). The dominance of diatom abundance in the tropical upwelling region (Table 4) was very similar to the coastal upwelling region (Wang et al., 2016). In addition, another significant result of this survey is the rise in dominant species abundance near 5°N, where the Halmahera Eddy (HE) and Mindanao Eddy (ME) are located (Fig. 4d). Phytoplankton primary production and growth could be stimulated by eddies in oligotrophic oceans (Falkowski et al., 1991). The anticyclonic HE and cyclonic ME may be the main reasons why dominant species abundance was high near 5°N.
Our results demonstrated the simultaneous occurrence of upwelling-induced nutrient supply and phytoplankton abundance (Table 4; Yuan et al., unpublished). Phytoplankton responded rapidly to the upwelling water that was nutrient-rich and highly saline (Feng, 1991; Kudela et al., 1997; Monteiro and Largier, 1999; Wang et al., 2016). Furthermore, the nutrients supplied by the upwelling contributed to the changes in phytoplankton abundance, biomass, and community structure (Feng, 1991; Irwin et al., 2006; Hong et al., 2011; Bode et al., 2015; Czerny et al., 2016; Wang et al., 2016). Our survey results suggested that epipelagic nutrient levels in the NECC station samples were high (Yuan et al., unpublished). Previous studies have shown that both the subtropical North Pacific gyre and the tropical western equatorial Pacific were low-nitrogen and low-phosphorus regions (Karl et al., 1997; Karl and Tien, 1997; Le Borgne and Rodier, 1997), which meant that the growth of phytoplankton was highly controlled by the nitrogen and phosphorus content. Our survey results also indicated that the marine diatoms T. longissimi, P. noctiluca, and C. frauenfeldianum, showed strong positive correlations with nitrogen and phosphorus (Fig. 5a). High nutrient water streams from the coastal MC and NGCC could also benefit the NECC region along with upwelling effects on nutrient supply (Christian et al., 2004; Higgins et al., 2006; Harsono et al., 2014). Many studies have reported that in marine environments, diatoms were dominant in the nutrient-rich nearshore and well-mixed waters (Pingree et al., 1978; Levasseur et al., 1984). In this study, phytoplankton, especially diatom abundance, was very abundant where the upwelling regions corresponded to regions with high nitrogen and phosphorus levels.
5 CONCLUSIONThis study reveals the spatial variability of phytoplankton communities with regards to composition, abundance, and biomass, and their relationships with physical oceanographic processes (currents, eddies, and fronts) and environmental factors (temperature, salinity, and nutrients) in the Western Boundary Currents. The environmental factors play a major role in distributing T. thiebautii filaments to the STCC region and the Luzon Strait. The impact of physical oceanographic processes (currents, eddies, and fronts) on phytoplankton cell abundance and biomass is strong. These results show that the current-driven upwellings and eddies increase phytoplankton abundance and biomass. Our data also show that the Western Boundary Currents are oligotrophic and complex environments and thus deserves special attention in future studies.
6 ACKNOWLEDGEMENTWe are grateful to YU Fei for the temperature and salinity data, ZHANG Wenjing for assisting with the chlorophyll concentration analyses and YUAN Yongquan for the nutrients data. We thank the captain and crew of R/V KeXue for their technical and logistical support.
| Ahmed A, Kurian S, Gauns M, Chndrasekhararao A V, Mulla A, Naik B, Naik H, Naqvi S W A, 2016. Spatial variability in phytoplankton community structure along the eastern Arabian Sea during the onset of south-west monsoon. Continental Shelf Research, 119: 30–39. Doi: 10.1016/j.csr.2016.03.005 |
| Angara E V, Rillon G S, Carmona M L, Ferreras J E M, Vallejo M I, Amper A C G G, Lacuna M L D G, 2013. Diversity and abundance of phytoplankton in Casiguran waters, Aurora Province, Central Luzon, Northern Philippines. AACL Bioflux, 6(4): 358–377. |
| Aurivillius C W S. 1898. Vergleichende thiergeographische Untersuchungen über die Plankton-Fauna des Skageraks in den Jahren 1893-1897. K. Svenska Vet. Akad. Handl., 30: 3. http://www.researchgate.net/publication/313199795_Vergleichende_thiergeographische_untersuchungen_uber_die_Plankton-Fauna_des_skageraks_in_den_jahren_1893-97 |
| Bibby T S, Moore C M, 2011. Silicate:nitrate ratios of upwelled waters control the phytoplankton community sustained by mesoscale eddies in sub-tropical North Atlantic and Pacific. Biogeosciences, 8(3): 657–666. Doi: 10.5194/bg-8-657-2011 |
| Bode A, Estévez M G, Varela M, Vilar J A, 2015. Annual trend patterns of phytoplankton species abundance belie homogeneous taxonomical group responses to climate in the NE Atlantic upwelling. Marine Environmental Research, 110: 81–91. Doi: 10.1016/j.marenvres.2015.07.017 |
| Breitbarth E, Oschlies A, LaRoche J, 2007. Physiological constraints on the global distribution of Trichodesmium-effect of temperature on diazotrophy. Biogeosciences, 4(1): 53–61. Doi: 10.5194/bg-4-53-2007 |
| Brown J S, Werner D, 1978. The biology of diatoms. Botanical Monographs, 13: 626. |
| Budaev S V, 2010. Using principal components and factor analysis in animal behaviour research:caveats and guidelines. Ethology, 116(5): 472–480. Doi: 10.1111/eth.2010.116.issue-5 |
| Capone D G, Zehr J P, Paerl H W, Bergman B, Carpenter E J, 1997. Trichodesmium, a globally significant marine cyanobacterium. Science, 276(5316): 1 221–1 229. Doi: 10.1126/science.276.5316.1221 |
| Carney R L, Seymour J R, Westhorpe D, Mitrovic S M, 2016. Lotic bacterioplankton and phytoplankton community changes under dissolved organic-carbon amendment:evidence for competition for nutrients. Marine and Freshwater Research, 67(9): 1 362–1 373. Doi: 10.1071/MF15372 |
| Carpenter E J, Janson S, 2000. Intracellular cyanobacterial symbionts in the marine diatom Climacodium frauenfeldianum (Bacillariophyceae). Journal of Phycology, 36(3): 540–544. Doi: 10.1046/j.1529-8817.2000.99163.x |
| Carpenter E J, 1983. Physiology and ecology of marine plankton Oscillatoria (Trichodesmium). Marine Biology Letters, 4: 69–85. |
| Chen Y L L, Chen H Y, Lin Y H, 2003. Distribution and downward flux of Trichodesmium in the South China Sea as influenced by the transport from the Kuroshio Current. Marine Ecology Progress Series, 259: 47–57. Doi: 10.3354/meps259047 |
| Chesson P, 2000. Mechanisms of maintenance of species diversity. Annual review of Ecology and Systematics, 31: 343–366. Doi: 10.1146/annurev.ecolsys.31.1.343 |
| Christian J R, Murtugudde R, Ballabrera-Poy J, McClain C R, 2004. A ribbon of dark water:phytoplankton blooms in the meanders of the pacific north equatorial countercurrent. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 51(1-3): 209–228. Doi: 10.1016/j.dsr2.2003.06.002 |
| Cleve P T. 1897. A Treatise on the Phytoplankton of the Atlantic and Its Tributaries and on the Periodical Changes of the Plankton of Skagerak. Upsala Nya Tidnings Aktiebolags Tryckeri, Upsala. 27p. http://agris.fao.org/agris-search/search.do?recordID=US201300223868 |
| Czerny J M S, Hauss H, Löscher C R, Riebesell U, 2016. Dissolved N:P ratio changes in the eastern tropical North Atlantic:effect on phytoplankton growth and community structure. Marine Ecology Progress Series, 545: 49–62. Doi: 10.3354/meps11600 |
| Everett J D, Baird M E, Roughan M, Suthers I M, Doblin M A, 2014. Relative impact of seasonal and oceanographic drivers on surface chlorophyll a along a Western Boundary Current. Progress in Oceanography, 120: 340–351. Doi: 10.1016/j.pocean.2013.10.016 |
| Falkowski P G, Ziemann D, Kolber Z, Bienfang P K, 1991. Role of eddy pumping in enhancing primary production in the ocean. Nature, 352(6330): 55–58. Doi: 10.1038/352055a0 |
| Feng J F. 1991. Composition and distribution of species on phytoplankton in Minnan-Taiwan bank fishing ground. In: Minnan-Taiwan Bank Fishing Ground Upwelling Ecosystem Study. Science Press, Beijing, China. p. 397-406. |
| Gomà J, Rimet F, Cambra J, Hoffmann L, Ector L, 2005. Diatom communities and water quality assessment in Mountain Rivers of the upper Segre basin (La Cerdanya, Oriental Pyrenees). Hydrobiologia, 551(1): 209–225. Doi: 10.1007/s10750-005-4462-1 |
| Hallegraeff G M, Jeffrey S W, 1984. Tropical phytoplankton species and pigments of continental shelf waters of north and north-west Australia. Marine ecology Progress Series, 20: 59–74. Doi: 10.3354/meps020059 |
| Harsono G, Atmadipoera A S, Syamsudin F, Manurung D, Mulyono S B, 2014. Halmahera Eddy features observed from multisensor satellite oceanography. Asian Journal of Scientific Research, 7(4): 571–580. Doi: 10.3923/ajsr.2014.571.580 |
| Hashihama F, Furuya K, Kitajima S, Takeda S, Takemura T, Kanda J, 2009. Macro-scale exhaustion of surface phosphate by dinitrogen fixation in the western North Pacific. Geophysical Research Letters, 36(3): L03610. |
| Hasle G R, Semina H J, 1987. The marine planktonic diatoms Thalassiothrix longissima and Thalassiothrix antarctica with comments on Thalassionema spp. and Synedra reinboldii. Diatom Research, 2(2): 175–192. Doi: 10.1080/0269249X.1987.9704996 |
| Higgins H W, Mackey D J, Clementson L, 2006. Phytoplankton distribution in the Bismarck Sea north of Papua New Guinea:the effect of the Sepik River outflow. Deep Sea Research Part Ⅰ:Oceanographic Research Papers, 53(11): 1 845–1 863. Doi: 10.1016/j.dsr.2006.09.001 |
| Hong H S, Wang Y J, Wang D Z, 2011. Response of phytoplankton to nitrogen addition in the Taiwan strait upwelling region:nitrate reductase and glutamine synthetase activities. Continental Shelf Research, 31(6): S57–S66. Doi: 10.1016/j.csr.2011.01.018 |
| Hood R R, Coles V J, Capone D G, 2004. Modeling the distribution of Trichodesmium and nitrogen fixation in the Atlantic Ocean. Journal of Geophysical Research:Oceans, 109(C6): C06006. |
| Hu D X, Wu L X, Cai W J, Gupta A S, Ganachaud A, Qiu B, Gordon A L, Lin X P, Chen Z H, Hu S J, Wang G J, Wang Q Y, Sprintall J, Qu T D, Kashino Y, Wang F, Kessler W S, 2015. Pacific western boundary currents and their roles in climate. Nature, 522(7556): 299–308. Doi: 10.1038/nature14504 |
| Hu J Y, Kawamura H, Hong H S, Qi Y Q, 2000. A review on the currents in the South China Sea:seasonal circulation, South China Sea warm current and Kuroshio intrusion. Journal of Oceanography, 56(6): 607–624. Doi: 10.1023/A:1011117531252 |
| Huang B Q, Hu J, Xu H Z, Cao Z R, Wang D X, 2010. Phytoplankton community at warm eddies in the northern South China Sea in winter 2003/2004. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 57(19-20): 1 792–1 798. Doi: 10.1016/j.dsr2.2010.04.005 |
| Irwin A J, Finkel Z V, Schofield O M E, Falkowski P G, 2006. Scaling-up from nutrient physiology to the size-structure of phytoplankton communities. Journal of Plankton Research, 28(5): 459–471. Doi: 10.1093/plankt/fbi148 |
| Jiang Y J, He W, Liu W X, Qin N, Ouyang H L, Wang Q M, Kong X Z, He Q S, Yang C, Yang B, Xu F L, 2014. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu). Ecological Indicators, 40: 58–67. Doi: 10.1016/j.ecolind.2014.01.006 |
| Jiang Z B, Zeng J N, Chen J F, Chen Q Z, Zhang D S, Yan X J, 2015. Diazotrophic cyanobacterium Trichodesmium spp. in China marginal seas:comparison with other global seas. Acta Ecologica Sinica, 35(2): 37–45. Doi: 10.1016/j.chnaes.2015.01.003 |
| Karl D M, Tien G, 1997. Temporal variability in dissolved phosphorus concentrations in the subtropical North Pacific Ocean. Marine Chemistry, 56(1-2): 77–96. Doi: 10.1016/S0304-4203(96)00081-3 |
| Karl D, Letelier R, Tupas L, Dore J, Christian J, Hebel D, 1997. The role of nitrogen fixation in biogeochemical cycling in the subtropical North Pacific Ocean. Nature, 388(6642): 533–538. Doi: 10.1038/41474 |
| Kimura S, Tsukamoto K, 2006. The salinity front in the North Equatorial Current:a landmark for the spawning migration of the Japanese eel (Anguilla japonica) related to the stock recruitment. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 53(3-4): 315–325. Doi: 10.1016/j.dsr2.2006.01.009 |
| Kitajima S, Furuya K, Hashihama F, Takeda S, Kanda J, 2009. Latitudinal distribution of diazotrophs and their nitrogen fixation in the tropical and subtropical western North Pacific. Limnology and Oceanography, 54(2): 537–547. Doi: 10.4319/lo.2009.54.2.0537 |
| Koslow J A, Pesant S, Feng M, Pearce A, Fearns P, Moore T, Matear R, Waite A, 2008. The effect of the Leeuwin Current on phytoplankton biomass and production off Southwestern Australia. Journal of Geophysical Research:Oceans, 113(C7): C07050. |
| Kudela R M, Cochlan W P, Dugdale R C, 1997. Carbon and nitrogen uptake response to light by phytoplankton during an upwelling event. Journal of Plankton Research, 19(5): 609–630. Doi: 10.1093/plankt/19.5.609 |
| Lacuna-Richman C, 2003. Ethnicity and the utilization of nonwood forest products:findings from three Philippine villages. Silva Fennica, 37(1): 129–148. |
| Langlois R J, Hümmer D, LaRoche J, 2008. Abundances and distributions of the dominant nifH phylotypes in the Northern Atlantic Ocean. Applied and Environmental Microbiology, 74(6): 1 922–1 931. Doi: 10.1128/AEM.01720-07 |
| Le Borgne R, Barber R T, Delcroix T, Inoue H Y, Mackey D J, Rodier M, 2002. Pacific warm pool and divergence:temporal and zonal variations on the equator and their effects on the biological pump. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 49(13-14): 2 471–2 512. Doi: 10.1016/S0967-0645(02)00045-0 |
| Le Borgne R, Rodier M, 1997. Net zooplankton and the biological pump:a comparison between the oligotrophic and mesotrophic equatorial Pacific. Deep Sea Research Part Ⅱ:Topical Studies in Oceanography, 44(9-10): 2 003–2 023. Doi: 10.1016/S0967-0645(97)00034-9 |
| Levasseur M, Therriault J C, Legendre L, 1984. Hierarchical control of phytoplankton succession by physical factors. Marine ecology Progress Series, 19: 211–222. Doi: 10.3354/meps019211 |
| Lipschultz F, Owens N J P, 1996. An assessment of nitrogen fixation as a source of nitrogen to the North Atlantic Ocean. Biogeochemistry, 35(1): 261–274. Doi: 10.1007/BF02179830 |
| Loder J W, Boicourt W C, Simpson J H. 1998. Western ocean boundary shelves, coastal segment (W), in The Sea. In: Brink K H, Robinson A R eds. The Global Coastal Ocean: Regional Studies and Syntheses. John Wiley, New York, USA. 11: 3-27. |
| Lorenzen C J, 1966. A method for the continuous measurement of in vivo chlorophyll concentration. Deep Sea Research and Oceanographic Abstracts, 13(2): 223–227. Doi: 10.1016/0011-7471(66)91102-8 |
| Lu G Y, Song X X, Yu Z M, Cao X H, Yuan Y Q, 2015. Environmental effects of modified clay flocculation on Alexandrium tamarense and paralytic shellfish poisoning toxins (PSTs). Chemosphere, 127: 188–194. Doi: 10.1016/j.chemosphere.2015.01.039 |
| Lukas R, Firing E, Hacker P, Richardson P L, Collins C A, Fine R, Gammon R, 1991. Observations of the Mindanao current during the western equatorial pacific ocean circulation study. Journal of Geophysical Research:Oceans, 96(C4): 7 089–7 104. Doi: 10.1029/91JC00062 |
| Moita M T, Pazos Y, Rocha C, Nolasco R, Oliveira P B, 2016. Toward predicting Dinophysis blooms off NW Iberia:a decade of events. Harmful Algae, 53: 17–32. Doi: 10.1016/j.hal.2015.12.002 |
| Monteiro P M S, Largier J L, 1999. Thermal stratification in Saldanha Bay (South Africa) and subtidal, density-driven exchange with the coastal waters of the Benguela upwelling system. Estuarine, Coastal and Shelf Science, 49(6): 877–890. Doi: 10.1006/ecss.1999.0550 |
| Okazaki Y, Nakata H, Kimura S, 2002. Effects of frontal eddies on the distribution and food availability of anchovy larvae in the Kuroshio Extension. Marine and Freshwater Research, 53(2): 403–410. Doi: 10.1071/MF01115 |
| Omura T, Iwataki M, Borja V M, Takayama H, Fukuyo Y. 2012. Marine Phytoplankton of the Western Pacific. Kouseisha Kouseikaku Co., Ltd., Tokyo, Japan. p. 25-148. |
| Pennington J T, Mahoney K L, Kuwahara V S, Kolber D D, Calienes R, Chavez F P, 2006. Primary production in the eastern tropical Pacific:a review. Progress in Oceanography, 69(2-4): 285–317. Doi: 10.1016/j.pocean.2006.03.012 |
| Pingree R D, Holligan P M, Mardell G T, 1978. The effects of vertical stability on phytoplankton distributions in the summer on the northwest European Shelf. Deep Sea Research, 25(11): 1 011–1 016. Doi: 10.1016/0146-6291(78)90584-2 |
| Reid P C, Surey-Gent S C, Hunt H G, Durrant A E, 1992. Thalassiothrix longissima, a possible oceanic indicator species in the North Sea. In ICES Mar. Sci. Symp, 195: 268–277. |
| Rivkin R B, Seliger H H, Swift E, Biggley W H, 1982. Lightshade adaptation by the oceanic dinoflagellates Pyrocystis noctiluca and P. fusiformis. Marine Biology, 68(2): 181–191. Doi: 10.1007/BF00397605 |
| Rodríguez J M, Hernández-León S, Barton E D, 1999. Mesoscale distribution of fish larvae in relation to an upwelling filament off Northwest Africa. Deep Sea Research Part Ⅰ:Oceanographic Research Papers, 46(11): 1 969–1 984. Doi: 10.1016/S0967-0637(99)00036-9 |
| Salomoni S E, Rocha O, Callegaro V L, Lobo E A, 2006. Epilithic diatoms as indicators of water quality in the Gravataí river, Rio Grande do Sul, Brazil. Hydrobiologia, 559(1): 233–246. Doi: 10.1007/s10750-005-9012-3 |
| Sañudo-Wilhelmy S A, Kustka A B, Gobler C J, Hutchins D A, Yang M, Lwiza K, Burns J, Capone D G, Raven J A, Carpenter E J, 2001. Phosphorus limitation of nitrogen fixation by Trichodesmium in the central Atlantic Ocean. Nature, 411(6833): 66–69. Doi: 10.1038/35075041 |
| Shaw P T, Chao S Y, 1994. Surface circulation in the South China Sea. Deep Sea Research Part Ⅰ:Oceanographic Research Papers, 41(11-12): 1 663–1 683. Doi: 10.1016/0967-0637(94)90067-1 |
| Shiozaki T, Kodama T, Kitajima S, Sato M, Furuya K, 2013. Advective transport of diazotrophs and importance of their nitrogen fixation on new and primary production in the western Pacific warm pool. Limnology and Oceanography, 58(1): 49–60. Doi: 10.4319/lo.2013.58.1.0049 |
| Shmida A, Wilson M V, 1985. Biological determinants of species diversity. Journal of Biogeography, 12(1): 1–20. Doi: 10.2307/2845026 |
| Stal L J, Severin I, Bolhuis H. 2010. The ecology of nitrogen fixation in cyanobacterial mats. In: Hallenbeck P. Recent Advances in Phototrophic Prokaryotes. Springer, New York, USA. p. 31-45. http://link.springer.com/10.1007/978-1-4419-1528-3_3 |
| Stal L J, 2009. Is the distribution of nitrogen-fixing cyanobacteria in the oceans related to temperature?. Environmental Microbiology, 11(7): 1 632–1 645. Doi: 10.1111/emi.2009.11.issue-7 |
| Stevenson R J, Pan Y D, van Dam H. 1999. Assessing environmental conditions in rivers and streams with diatoms. In: Smol J P, Stoermer E F eds. The Diatoms: Applications for the Environmental and earth Sciences. 2nd edn. Cambridge University Press, Cambridge, Britain. 4p. |
| Tang D L, Ni I H, Kester D R, Müller-Karger F E, 1999. Remote sensing observations of winter phytoplankton blooms southwest of the Luzon Strait in the South China Sea. Marine Ecology-Progress Series, 191: 43–51. Doi: 10.3354/meps191043 |
| Taylor F J R. 1973. General features of dinoflagellate material collected by the "Anton Bruun" during the International Indian Ocean Expedition. In: Zeitzschel B, Gerlach S A eds. The Biology of the Indian Ocean. Springer, Berlin Heidelberg, Germany. p. 155-169. https://link.springer.com/chapter/10.1007%2F978-3-642-65468-8_11 |
| Thorson G, 1950. Reproductive and larval ecology of marine bottom invertebrates. Biological Reviews, 25(1): 1–45. Doi: 10.1111/brv.1950.25.issue-1 |
| Tilman D, Kilham S S, Kilham P, 1982. Phytoplankton community ecology:the role of limiting nutrients. Annual Review of Ecology and Systematics, 13: 349–372. Doi: 10.1146/annurev.es.13.110182.002025 |
| Tyrrell T, 1999. The relative influences of nitrogen and phosphorus on oceanic primary production. Nature, 400(6744): 525–531. Doi: 10.1038/22941 |
| Vaillancourt R D, Marra J, Seki M P, Parsons M L, Bidigare R R, 2003. Impact of a cyclonic eddy on phytoplankton community structure and photosynthetic competency in the subtropical North Pacific Ocean. Deep Sea Research Part Ⅰ:Oceanographic Research Papers, 50(7): 829–847. Doi: 10.1016/S0967-0637(03)00059-1 |
| Voss M, Croot P, Lochte K, Mills M, Peeken I, 2004. Patterns of nitrogen fixation along 10°N in the tropical Atlantic. Geophysical Research Letters, 31(23): L23S09. |
| Wang Y, Kang J H, Ye Y Y, Lin G M, Yang Q L, Lin M, 2016. Phytoplankton community and environmental correlates in a coastal upwelling zone along western Taiwan Strait. Journal of Marine Systems, 154: 252–263. Doi: 10.1016/j.jmarsys.2015.10.015 |
| Yang Q L, Lin G M, Lin M, Dai Y Y, 2002. Species Composition and distribution of phytoplankton in the china pioneer area of the northeast Pacific Ocean. Marine Science Bulletin, 21(4): 15–27. |
| Yang Y, Sun X X, Zhu M L, Luo X, Zheng S. 2016. Estimating the carbon biomass of marine net phytoplankton from abundance based on samples from China seas. Marine and Freshwater Research. https://doi.org/10.1071/mf15298. |
| Yentsch C S, Menzel D W, 1963. A method for the determination of phytoplankton chlorophyll and phaeophytin by fluorescence. Deep Sea Research and Oceanographic Abstracts, 10(3): 221–231. Doi: 10.1016/0011-7471(63)90358-9 |
| Zehr J P, Ward B B, 2002. Nitrogen cycling in the ocean:new perspectives on processes and paradigms. Applied and Environmental Microbiology, 68(3): 1 015–1 024. Doi: 10.1128/AEM.68.3.1015-1024.2002 |