 2018, Vol. 36
2018, Vol. 36Institute of Oceanology, Chinese Academy of Sciences
Article Information
- Bhushan Kumar SHARMA, Shaikhul Islam KHAN, Sumita SHARMA
- Biodiverse rotifer assemblage (Rotifera: Eurotatoria) of floodplain lakes of the Brahmaputra basin of lower Assam, northeast India: composition and ecosystem diversity
- Chinese Journal of Oceanology and Limnology, 36(2): 362-375
- http://dx.doi.org/10.1007/s00343-017-6251-x
Article History
- Received Sep. 27, 2016
- accepted in principle Nov. 20, 2016
- accepted for publication Dec. 15, 2018




The floodplain lakes form an integral component of the major riverine systems world-wide. These wetlands attract attention as a lucrative source of inland fisheries and for rich aquatic biodiversity. These generalizations hold valid for the floodplain lakes (beels) of the Brahmaputra river basin of NEI (Sugunan, 1997; Sharma and Sharma, 2008, 2013) and those of lower Assam in particular. The latter are subjected to unplanned fishing with removal of aquatic vegetation thus altering their ecotone character. Referring to biodiversity value, Segers et al. (1993) firstly hypothesized tropical and subtropical floodplain lakes to be globally interesting habitats for the rotifer diversity. Sharma and Sharma(2008, 2014a, 2014b) extended this hypothesis to the Brahmaputra floodplains of the Indian sub-region based on analysis of extensive plankton and semiplankton collections. The noteworthy reports of 171 and 162 rotifer species from Deepor beel (Sharma and Sharma, 2015a) and Loktak Lake (Sharma et al., 2016)—two Ramsar sites of India and important NEI wetlands, respectively hold distinction as one of the richest rotifer habitats. Rotifera, an integral link of aquatic food-webs, are also known to be useful bioindicators and thus useful to depict trophic status of water bodies (Sládeček, 1983).
Realizing the rotifer biodiversity value of the Brahmaputra basin beels vis-à-vis the role of sampling, use of these metazoans to assess trophic status and to characterize habitat variations, we undertook limnological reconnaissance of four beels of lower Assam. Our study resulted in the detailed analysis of Rotifera assemblages with species-rich inventory of the taxon and interesting features. The nature and composition of the rotifer fauna are discussed with reference to biodiversity, distribution and ecosystem diversity, and various interesting rotifers are illustrated. Remarks are made on monthly richness variations, community similarities and on individual as well as cumulative influence of abiotic factors. We propose L/B quotient based on Lecane: Brachionus species ratios to depict habitat variations and use B/T quotient (Sládeček, 1983) to comment on trophic status of the sampled wetlands.
2 MATERIAL AND METHODThis study is a part of limnological survey of four floodplain lakes (26o17′08.7″–26o30′22.2″N; 90o52′12.8″–91o02′43.1″E; altitude: 37.9–54.6 m ASL) namely Fingua, Baria, Sorbhog and Balaisuti beels of the Barpeta district of lower Assam, NEI (Fig. 1a, b) undertaken during November 2011– October 2013. Eichhornia crassipes (Mart.) Solms, Hydrilla verticillata (L. f.) Royle, Pistia stratiotes Linnaeus., Salvinia cucullata Roxb., Azolla sp. and Trapa natans L. commonly occurred while Nymphoides sp., Ipomoea sp., Lemna sp., Hygroryza aristata (Retz.) Nees, Oryza rufipogon Griff. and Nelumbo sp. are other aquatic plants of these wetlands. These beels experienced habitat disturbances during intensive fishing from November-February with removal of aquatic vegetation and varying extent of unplanned fishing from January–October during each year.

|
| Figure 1 Map of India showing Northeast India (NEI) and Assam state (green) (a); map of Assam indicating Barpeta district (b) |
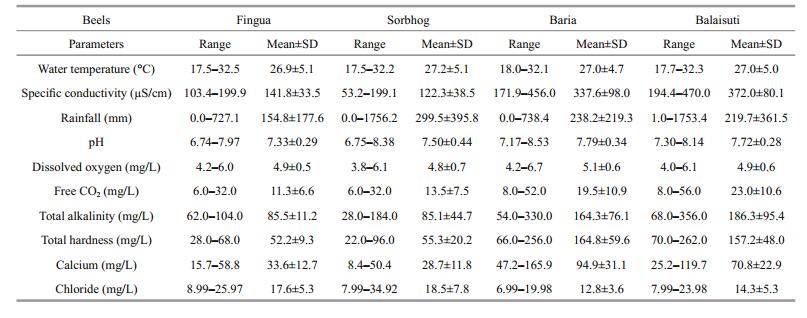
Water samples were collected monthly from four beels and analyzed for ten abiotic factors. Water temperature, specific conductivity and pH were recorded in the field using relevant probes; Winkler's method was used for estimation of dissolved oxygen and Greenberg (1992) was followed to analyze total alkalinity, total hardness, calcium and chloride. The rainfall data was obtained from local meteorological sources. The qualitative plankton samples were collected monthly from Fingua, Baria, Sorbhog and Balaisuti beels by towing a nylobolt plankton net (# 50 μm) and were preserved in 5% formalin. Plankton samples were screened, individual rotifers were isolated and mounted in Polyvinyl alcohollactophenol, and were observed with Leica (DM 1000) stereoscopic phase contrast microscope using an image analyzer. Interesting rotifers were illustrated and measurements were given in micrometers (μm). The rotifers taxa were identified following Koste (1978), Segers (1995), Sharma (1998), Sharma and Sharma(1999, 2008, 2013). The community similarities were calculated vide Sørensen's index and SPSS (Version 20) was used for hierarchical cluster analysis. Two-way ANOVA was used to ascertain significance of richness variations. The relationships between abiotic factors and richness were determined by Pearson's correlation coefficients (R). The canonical correspondence analysis was done using XLSTAT (2015) to analyze cumulative influence of 10 abiotic parameters namely water temperature, rainfall, pH, specific conductivity, dissolved oxygen, free carbon dioxide, total alkalinity, total hardness, calcium and chloride on the rotifer richness.
We propose L/B quotient, based on Lecane: Brachionus species ratios, to characterize habitat changes of the sampled beels. As per our results, L/B quotient values up to 1.0; 1.0–3.0; and above 3.0 categorized limnetic, semi-limnetic and littoralperiphytic habitat nature, respectively. Comments on trophic status of the beels are based on B/T quotient based on Brachionus: Trichocerca species ratios following Sládeček (1983).
3 RESULTThe variations (ranges, mean±SD) of ten abiotic factors are presented in Table 1. Water temperature varied between (26.9±5.1)–(27.2±5.1)℃, pH between (7.33±0.29)–(7.79±0.34) and specific conductivity between (122.3±38.5)–(372.0±80.1) μS/cm while dissolved oxygen and free carbon dioxide fluctuated between (4.0±0.5)–(5.1±0.6) mg/L and (11.3±6.6)– (23.0±10.6) mg/L, respectively amongst the sampled beels. The monthly rainfall, total alkalinity, total hardness, calcium and chloride ranged between (154.8±177.6)–(299.5±395.8) mm, (85.1±44.7– (186.3±95.4) mg/L; (52.2±9.3)–(164.8±59.6) mg/L; (28.7±11.8)–(94.9±31.1) mg/L and (12.8±3.6)– (18.5±7.8) mg/L, respectively.
A total of 160 species belonging to 19 families and 35 genera are observed from four beels of Barpeta, lower Assam (Appendix Ⅰ). Cephalodella trigona (Fig. 2a) is a new record to the Oriental region; Lecane bifastigata (Fig. 2b) is new to NEI; and Trichocerca hollaerti (Fig. 2c), T. maior (Fig. 2d) and Dissotrocha aculeata are new records from Assam. Besides, we recorded the Oriental Brachionus donneri (Fig. 2e) and Lecane superaculeata (Fig. 2f), and the Australasian Notommata spinata (Fig. 2g) while Lecane dorysimilis (Fig. 2h), L. hastata (Fig. 2i), L. rhenana (Fig. 2j) and Mytilina michelangellii (Fig. 2k) are other interesting species.

|
| Figure 2 a. Cephalodella trigona (Rousselet) (lateral view); b. Lecane bifastigata Hauer (dorsal view); c. Trichocerca hollaerti De Smet (lateral view); d. Trichocerca maior Hauer (lateral view); e. Brachionus donneri Brehm (ventral view); f. Lecane superaculeata Sanoamuang & Segers (ventral view); g. Notommata spinata Koste & Shiel (ventral view); h. Fig. 19, Lecane dorysimilis Trinh Dang, Segers & Sanoamuang (dorsal view); i. L. rhenana Hauer (ventral view); j. Lecane hastata (Murray (ventral view); k. Mytilina michelangellii Reid & Turner (lateral view) |
Total rotifer richness in different beels ranged between 99 (Balaisuti)–124 (Sorbhog) species during the study period (Fig. 3) with mean value of 110±10 species; it ranged between 65 (Baria) -92 (Sorbhog) and 82(Balaisuti) -108 (Sorbhog) species and recorded 67.8%–74.4% and 70.3%–78.4% community similarities (S rensen's index) amongst beels during two years, respectively. The monthly richness (Figs. 4–7) ranged between 5–44 (23±11), 6–47 (23±10), 4–56 (29±13) and 7–46 (27±9) species, and the rotifer community similarities ranged between 0–63.2% and 4.6%–71.0%; 0–85.7% and 8.2%– 77.2%; 0–71.7% and 16.4%–54.5%; and 16.2%– 69.8% and 32.7%–68.9% in Fingua, Baria, Sorbhog and Balaisuti beels during two years, respectively. Annual hierarchical cluster analysis of Rotifera amongst beels is presented in Figs. 8–9. ANOVA registered significant richness variations amongst four beels (F3, 69=3.382, P=0.023 0) and significant monthly variations (F3, 23=4.627, P=3.902E-07) between them. Further, it registered significant annual richness variations in Baria (F1, 11=7.604, P=0.018 6), Sorbhog (F1, 11=5.167, P=0.037 1) and Balaisuti (F1, 11= 6.034, P=0.031 9) beels.

|
| Figure 3 Species richness of Rotifera of Barpeta beels |

|
| Figure 4 Monthly variations of Rotifera richness in Fingua beel (2011–2013) |

|
| Figure 5 Monthly variations of Rotifera richness in Baria beel (2011–2013) |

|
| Figure 6 Monthly variations of Rotifera richness in Sorbhog beel (2011–2013) |

|
| Figure 7 Monthly variations of Rotifera richness in Balaisuti beel (2011–2013) |

|
| Figure 8 The hierarchical cluster analysis of Rotifera assemblages (2011–2012) |

|
| Figure 9 The hierarchical cluster analysis of Rotifera assemblages (2012–2013) |
Lecanidae, Brachionidae, Lepadellidae and Trichocercidae indicated 46, 28, 22 and 15 species, respectively. Testudinellidae (9 species), Notommatidae (7 species), Euchlanidae (6 species) and Mytilinidae (6 species) are other species-rich families. Lecane (46 species), Brachionus (18 species), Lepadella (17 species) and Trichocerca (15 species) are important genera. Lecane / Brachionus (L/B) quotient indicated oscillating monthly variations (Fig. 11) with mean values of 3.7±3.3, 3.5±4.1, 4.1±5.1 and 4.5±2.7 in four beels, respectively. Sládeček's Brachionus / Trichocerca (B/T) quotient followed oscillating monthly variations in all beels (Fig. 10) with mean values of 1.6±1.5, 2.8±2.2, 2.8±2.1 and 2.2±1.3 in Fingua, Baria, Sorbhog and Balaisuti beels, respectively.Of ten abiotic factors, only total alkalinity recorded significant negative correlation with richness (R=-0.597, P=0.002 1) in Balaisuti beel. The canonical correspondence analysis (CCA) co-ordination biplots of Rotifera richness with ten abiotic parameters (Figs. 12–15) indicated cumulative influences of 88.15%, 80.72%, 94.70% and 92.04%, along two axes, in Fingua, Baria, Sorbhog and Balaisuti beels, respectively.

|
| Figure 10 Monthly variations of Brachionus / Trichocerca species ratios in beels (2011–2013) |

|
| Figure 11 Monthly variations of Lecane / Brachionus species ratios in beels (2011–2013) |

|
| Figure 12 CCA coordination biplot of Rotifera richness and abiotic factors of Fingua beel Abbreviations: Abiotic: Ca (Calcium), Cl (Chloride), Cond (conductivity), DO (dissolved oxygen), FCO (free carbon dioxide), TA (total alkalinity), TH (total hardness), pH (hydrogen-ion concentration), Rain (rainfall), Wt (water temperature). Biotic: Bra (Brachionidae), Br sp (Brachionus spp.), B/T (Brachionus/Trichocerca ratio), Lec (Lecanidae), L/B (Lecane/Brachionus ratio), RR (Rotifera richness), ZR (zooplankton richness). |

|
| Figure 13 CCA coordination biplot of Rotifera richness and abiotic factors of Baria beel Abbreviations: Abiotic: Ca (Calcium), Cl (Chloride), Cond (conductivity), DO (dissolved oxygen), FCO (free carbon dioxide), TA (total alkalinity), TH (total hardness), pH (hydrogen-ion concentration), Rain (rainfall), Wt (water temperature). Biotic: Bra (Brachionidae), Br sp (Brachionus spp.), B/T (Brachionus/Trichocerca ratio), Lec (Lecanidae), L/B (Lecane/Brachionus ratio), RR (Rotifera richness), ZR (zooplankton richness). |

|
| Figure 14 CCA coordination biplot of Rotifera richness and abiotic factors of Sorbhog beel Abbreviations: Abiotic: Ca (Calcium), Cl (Chloride), Cond (conductivity), DO (dissolved oxygen), FCO (free carbon dioxide), TA (total alkalinity), TH (total hardness), pH (hydrogen-ion concentration), Rain (rainfall), Wt (water temperature). Biotic: Bra (Brachionidae), Br sp (Brachionus spp.), B/T (Brachionus/Trichocerca ratio), Lec (Lecanidae), L/B (Lecane/Brachionus ratio), RR (Rotifera richness), ZR (zooplankton richness). |

|
| Figure 15 CCA coordination biplot of Rotifera richness and abiotic factors of Balaisuti beel Abbreviations: Abiotic: Ca (Calcium), Cl (Chloride), Cond (conductivity), DO (dissolved oxygen), FCO (free carbon dioxide), TA (total alkalinity), TH (total hardness), pH (hydrogen-ion concentration), Rain (rainfall), Wt (water temperature). Biotic: Bra (Brachionidae), Br sp (Brachionus spp.), B/T (Brachionus/Trichocerca ratio), Lec (Lecanidae), L/B (Lecane/Brachionus ratio), RR (Rotifera richness), ZR (zooplankton richness). |
We characterize Barpeta beels into two categories based on pH, specific conductivity, total alkalinity, total hardness and Calcium. Fingua and Sorbhog belonged to first category with circumneutral to slightly alkaline and moderately hard waters with low specific conductivity and calcium. Baria and Balaisuti beels with alkaline, hard and calcium rich waters with more than 2.5 times specific conductivity are grouped together. The second group differed from earlier reports on floodplains of Assam (Sharma and Sharma, 2008, 2014a; Sharma, 2014, 2016; Sharma et al., 2015) and Manipur (Sharma, 2009a, 2009b, 2011; Sharma et al., 2016) states of NEI.
A total of 160 rotifer species (S) observed from four beels is one of the richest assemblages of the taxon known from the Indian floodplains. This report is of biodiversity value as ~38.0% and ~57.0% of species known till date from India and NEI, respectively (BKS, unpublished). The biodiverse Rotifera is hypothesized to habitat diversity and environmental heterogeneity of the sampled wetlands. Our results supported hypothesis of Segers et al. (1993) indicating (sub) tropical floodplains to be the world's rotifer rich habitats. The diverse nature corroborated with the results from the floodplains of Argentina (de Paggi, 1993, 2001), Brazil (Bonecker et al., 1998) and Australia (Shiel et al., 1998). Further, 35 genera and 19 families recorded presently affirmed higher diversity of the taxon.
Cephalodella trigona is new to the Oriental Rotifera. The present report of this palaearctic species (Segers, 2007) extended its distribution to the Indian sub-region. Lecane bifastigata is new to NEI. This lecanid is known from Palaearctic and Oriental regions (Segers, 2007) and it is validly reported from India from West Bengal (Sharma, 1979; Sharma and Sharma, 2014c). Trichocerca hollaerti, T. maior and Dissotrocha aculeata are new records from Assam. The first two species are added to the Indian Rotifera from Mizoram (Sharma and Sharma, 2015b) and are also reported from Loktak Lake (a Ramsar site), Manipur (Sharma et al., 2016) while D. aculeata is reported from India from a wetland of Meghalaya (Sharma, 2016). Besides, our earlier report (Sharma and Khan, 2016) from these beels added Testudinella dendradena and Brachionus bennini as new to the Indian Rotifera and to the rotifer fauna of Assam, respectively.
Twenty-three globally interesting species formed an important fraction (14.4% of S) of biogeographic value. These included (ⅰ) the Australasian Brachionus dichotomus reductus, B. kostei and Notommata spinata; (ⅱ) the Oriental endemics: Brachionus donneri, Keratella edmondsoni, Lecane blachei, L. bulla diabolica, L. niwati, L. superaculeata and Filinia camasecla; (ⅲ) the paleotropical species namely Keratella javana, Lepadella discoidea, L. vandenbrandei, Lecane lateralis, L. simonneae, L. unguitata, Testudinella brevicaudata, T. greeni and Trichocerca hollaerti; the palaeotropical Cephalodella trigona; Holarctic Trichocerca maior; the IndoChinese Lecane dorysimilis and the cosmo(sub) tropical Brachionus durgae. Of these, comments on 15 species are made earlier by Sharma and Khan (2016).
Brachionus diversicornis, B. forficula, Keratella javana, Lecane lateralis, L. simonneae, L. unguitata, Lepadella discoidea, L. vandenbrandei, Testudinella brevicaudata and T. greeni are 'Eastern hemisphere elements' (vide Segers, 2001). Brachionus dichotomus reductus, B. kostei, Keratella javana, Lepadella benjamini, L. vandenbrandei, Lecane bifastigata, L. blachei, L. dorysimilis, L. niwati, L. rhenana, L. rhytida, L. superaculeata, Monommata grandis, Notommata spinata, Testudinella amphora, T. brevicaudata, T. dendradena, T. greeni, T. tridentata, Trichocerca bidens and T. hollaerti are species of regional value in the Indian sub-region with distribution restricted till date to NEI. In addition, Brachionus bennini, B. donneri, B. leydigii, Filinia camasecla, F. saltator, Keratella edmondsoni, K. tecta, Lacinularia flosculosa, Lecane aspasia, L. bifurca, L. bulla diabolica, L. haliclysta, L. lateralis, L. pusilla, L. simonneae, L. undulata, Lepadella biloba, L. costatoides, L. dactyliseta, L. discoidea, L. quinquecostata, L. triba, Macrochaetus longipes, Mytilina michelangellii, Platyias leloupi, Trichocerca insignis, T. maior, T. scipio and T. tigris are species with restricted distribution in the Indian sub-continent.
The overall rotifer richness (S) is lower than 207, 218 and 252 species known from the floodplains of Africa (Segers et al., 1993), South America (Bonecker et al., 1998) and Australia (Shiel et al., 1998), respectively. This comparison needed caution as our inventory included only planktonic and semiplanktonic taxa. Total richness compared well the report of 164 species from 15 beels of lower Assam (Sharma, 2005); 162 species from Loktak Lake, Manipur (Sharma et al., 2016). On the other hand, rotifera of lower Assam beels is diverse than the reports of 144 species from the Majuli floodplains, upper Assam (Sharma et al., 2015), 124 species from seven beels of lower Assam (Sharma and Sharma, 2001) and 110 species (Arora and Mehra, 2003) from the backwaters of the river Yamuna at Delhi.
Our collections indicated diverse Lecanidae (28.7% of S) and the collective importance (40.0% of S) of Brachionidae > Lepadellidae > Trichocercidae. These four Eurotatoria families exhibited biodiversity interest (69.4% of S) concurrent with the reports from Deepor beel (Sharma and Sharma, 2015a) and Loktak Lake (Sharma et al., 2016). The notable richness of Brachionidae (17.5% of S) and of Brachionus spp. (11.2%) concurred with the report from Deepor beel (Sharma and Sharma, 2015a) but it contrasted than brachionid paucity reports of Sharma(2009a, 2009b, 2014) and Sharma et al.(2015, 2016) from NEI. High brachionid diversity in Barpeta beels is attributed to their alkaline hard waters and to limnetic conditions created due to mass removal of aquatic vegetation during fishing. The species of littoral-periphytic Lecane > Lepadella > Trichocerca, together, comprised a notable fraction (~49.0% of S). The stated importance supported hypothesis of Green (2003) on the possibility of assemblage rules for the periphytic community. Further, this salient feature concurred with the reports from the floodplains of Brazil (Segers et al., 1993; Segers and Dumont, 1995), Thailand (Sanoamuang 1998), Bolivia (Segers et al., 1998) and South Africa (Green, 2003) and India (Sharma and Sharma, 2014a). The rich nature and common occurrence of 'tropic-centered genera' Lecane and Brachionus, more cosmopolitan species (~63.0% of S), and collective richness (~20% of S) of cosmotropical and pantropical species imparted 'tropical character' to Barpeta Rotifera. This generalization is concurrent with composition of several tropical rotifer faunas (Green, 1972; Fernando, 1980; Dussart et al., 1984; Segers, 1996, 2001).
We observed high overall rotifer richness in four beels (99–124, 110±10 species) with Sorbhog > Fingua > Baria > Balaisuti beels. The mean richness concurred with the reports of 114 species (de Paggi, 2001) from the Rio Pilcomayo National park (a Ramsar site), Argentina; 106 taxa from Thale-Noi Lake, Thailand (Segers and Pholpunthin, 1997); and 104 species from Laguana Bufeos, Bolivia (Segers et al., 1998). Rotifera of Barpeta beels is, however, more biodiverse than the previous report of 73 species from Fingua beel (Sharma, 2005) and from the reports of 85 species (Sharma and Sharma, 2012), 69–93 species (Sharma and Sharma, 2008) and 60–100 species (Sharma et al., 2015) from various beels of Assam. Our report also showed diverse nature than the records of 71 and 75 species from Utra and Waithou pats (Sharma, 2011) and 62–73 species from 15 pats of Manipur (Sharma, 2009b). Total richness indicated inter-annual variations with higher range and mean during second year. High community similarities (67.8%–74.4% and 70.3%–78.4%) noticed during two years, respectively reflected homogeneity in the rotifer species composition amongst beels. This generalization is also supported by occurrence of 41 species (25.6 % of S) in all beels. ANOVA registered significant richness variations amongst four beels and significant monthly variations between them. The hierarchical cluster indicated the rotifer affinity between Fingua and Baria beels while Sorbhog beel showed distinct divergence during both years.
Individual beels are characterized by heterogeneity in Rotifera composition as endorsed by wider monthly richness variations, lower mean values, and lower yet wider community similarities in majority of instances and lack of occurrence of any species throughout the study period in any beel. The rotifer heterogeneity in individual beels is hypothesized to habitat disturbance due to fishing and removal of weeds though specific observations are desired this with effect. ANOVA registered significant annual richness variations in Baria, Sorbhog and Balaisuti beels.
Lecane / Brachionus (L/B) quotient mean values affirmed general littoral-periphytic character of different beels but oscillating monthly variations yet depicted habitat changes. The latter is reflected by low L/B values during January–April and February– June in Fingua; December–March, August–February and May–June in Baria; December–April, July– August, November and March–May in Sorbhog; and January, October and December in Balaisuti beel during the study period. Nevertheless, L/B values > 6.0 and > 10.0, observed during certain months and indicating typical wetland character in all beels, are attributed to low Brachionus richness or lack of species of the genus, respectively. The results, though not indicative of definitive patterns, asserted usefulness of L/B quotient vis-à-vis habitat changes in the sampled beels.
Brachionus/Trichocerca quotient (vide Sládeček, 1983), an analogue of phytoplankton indices to establish trophic status of lentic or lotic waters, followed oscillating monthly variations in all beels while mean values indicated overall 'mesotrophic' nature of four beels. Nevertheless, B/T recorded values up to 1.0 in 18, nine, seven and eight months in Fingua, Baria, Sorbhog and Balaisuti beels, respectively while > 3.0 values are recorded in eight and nine instances in Baria and Sorbhog beels, respectively while Fingua and Balaisuti beels recorded 'eutrophic' character in three and two instances, respectively. Sladecek's quotient depicted trophic status variations in the sampled beels; the results ascertained reliability of use of this quotient under Indian conditions concurrent with the remarks of Sharma and Dudani (1992), and Sharma and Lyngskor (2003).
The lack of impact of individual abiotic factors on the rotifer richness in Barpeta beels is endorsed by only significant negative correlation of total alkalinity in Balaisuti beel. The canonical correspondence analysis affirmed higher cumulative influences (80.72%–94.70%) in different beels along two axes with higher values > 92.0% in Sorbhog and Balaisuti beels. Chloride, rainfall and total harness; rainfall, water temperature, total hardness; water temperature, chloride, total alkalinity; water temperature, free carbon dioxide and calcium influenced richness of Rotifera in Fingua, Baria, Sorbhog and Balaisuti beels, respectively in CCA bi-plots.
5 CONCLUSIONThe speciose and diverse Rotifer assemblage, taxa of global and regional distribution interest and Eastern Hemisphere species impart biodiversity importance to this study as well as value for meta-analysis of the rotifer diversity in the Indian floodplains. The richness variations amongst and within different beels vis-à-vis homogeneity or heterogeneity in species composition, oscillating monthly richness, cumulative influence of abiotic factors rather than of individual factors, generalizations and variations of B/T and L/B quotients are useful attributes of ecosystem diversity.
6 ACKNOWLEDGEMENTThe senior author (BKS) thanks the Head, Department of Zoology, North-Eastern Hill University, Shillong for laboratory facilities. The samples analyzed for this study were collected by SIK. The authors have no conflict of research interests. Finally, we sincerely thank our anonymous reviewers for their valuable comments and suggestions.
Appendix Ⅰ: Systematic list of Rotifera recorded from Beels of lower AssamPhylum: Rotifera
Class: Eurotatoria
Subclass: Monogononta
Order: Ploima
Family: Brachionidae
1. Anuraeopsis fissa (Gosse, 1851)
2. Brachionus ahlstromi Lindeman, 1939
3. B. angul aris Gosse, 1851
4. B. bennini Leissling, 1924 #
5. B. bidentatus Anderson, 1889
6. B. budapestinensis Daday, 1885
7. B. calyciflorus Pallas, 1766
8. B. caudatus Barrois & Daday, 1894
9. B. donneri Brehm, 1951
10. B. dichotomus reductus Koste & Shiel, 1980 #
11. B. diversicornis (Daday, 1883)
12. B. durgae Dhanapathi, 1974 #
13. B. falcatus Zacharias, 1898
14. B. forficula Wierzejski, 1891
15. B. kostei Shiel, 1983 #
16. B. leydigii Cohn, 1862
17. B. mirabilis Daday, 1897
18. B. quadridentatus Hermann, 1783
19. B. rubens Ehrenberg, 1838
20. Keratella cochlearis (Gosse, 1851)
21. K. edmondsoni Ahlstrom, 1943 #
22. K. javana Hauer, 1937 #
23. K. lenzi Hauer, 1953
24. K. tecta (Gosse, 1851)
25. K. tropica (Apstein, 1907)
26. Plationus patulus (O. F. Muller, 1786)
27. Platyias leloupi (Gillard, 1967)
28. P. quadricornis (Ehrenberg, 1832)
Family: Euchlanidae
29. Beauchampiella eudactylota (Gosse, 1886)
30. Dipleuchlanis propatula (Gosse, 1886)
31. Euchlanis dilatata Ehrenberg, 1832
32. E. incisa Carlin, 1939
33. E. triquetra Ehrenberg, 1838
34. Tripleuchlanis plicata (Levander, 1894)
Family: Mytilinidae
35. Lophocharis salpina (Ehrenberg, 1834)
36. Mytilina acanthophora Hauer, 1938
37. M. bisulcata (Lucks, 1912)
38. M. brevispina (Ehrenberg, 1830)
39. M. michelangellii Reid & Turner, 1988
40. M. ventralis (Ehrenberg, 1830)
Family: Trichotriidae
41. Macrochaetus collinsi (Gosse, 1867)
42. M. longipes Myers, 1934
43. M. sericus (Thorpe, 1893)
44. Trichotria tetractis (Ehrenberg, 1830)
Family: Lepadellidae
45. Colurella adriatica Ehrenberg, 1831
46. C. obtusa (Gosse, 1886)
47. C. sulcata (Stenroos, 1898)
48. C. uncinata (O. F. Muller, 1773)
49. Lepadella acuminata (Ehrenberg, 1834)
50. L. apsida Harring, 1916
51. L. benjamini Harring, 1916
52. L. biloba Hauer, 1958
53. L. costatoides Segers, 1992
54. L. dactyliseta (Stenroos, 1898)
55. L. discoidea Segers, 1993 #
56. L. ovalis (O. F. Muller, 1786)
57. L. patella (O. F. Muller, 1773)
58. L. quinquecostata (Lucks, 1912)
59. L. rhomboides (Gosse, 1886)
60. L. triba Myers, 1934
61. L. triptera Ehrenberg, 1832
62. L. vandenbrandei Gillard, 1952 #
63. L. (H.) apsicora Myers, 1934
64. L. (H.) ehrenbergi (Perty, 1850)
65. L. (H.) heterostyla (Murray, 1913)
66. Squatinella lamellaris (O. F. Müller, 1786)
Family: Lecanidae
67. Lecane aculeata (Jakubski, 1912)
68. L. arcuata (Bryce, 1891)
69. L. aspasia Myers, 1917
70. L. bifastigata Hauer, 1938 **
71. L. bifurca (Bryce, 1892)
72. L. blachei Berzins, 1973 #
73. L. bulla bulla (Gosse, 1851)
L. bulla diabolica (Hauer, 1936) #
74. L. closterocerca (Schmarda, 1859)
75. L. crepida Harring, 1914
76. L. curvicornis (Murray, 1913)
77. L. decipiens (Murray, 1913)
78. L. dorysimilis Trinh Dang, Segers & Sanoamuang, 2015
79. L. doryssa Harring, 1914
80. L. flexilis (Gosse, 1886)
81. L. furcata (Murray, 1913)
82. L. haliclysta Harring & Myers, 1926
83. L. hamata (Stokes, 1896)
84. L. hastata (Murray, 1913)
85. L. hornemanni (Ehrenberg, 1834)
86. L. inermis (Bryce, 1892)
87. L. inopinata Harring & Myers, 1926
88. L. lateralis Sharma, 1978 #
89. L. leontina (Turner, 1892)
90. L. ludwigii (Eckstein, 1883)
91. L. luna (Müller, 1776)
92. L. lunaris (Ehrenberg, 1832)
93. L. monostyla (Daday, 1897)
94. L. nitida (Murray, 1913)
95. L. niwati Segers, Kothetip & Sanoamuang, 2004 #
96. L. obtusa (Murray, 1913)
97. L. ohioensis (Herrick, 1885)
98. L. papuana (Murray, 1913)
99. L. ploenensis (Voigt, 1902)
100. L. pusilla Harring, 1914
101. L. pyriformis (Daday, 1905)
102. L. quadridentata (Ehrenberg, 1830)
103. L. rhenana Hauer, 1929
104. L. rhytida Harring & Myers, 1926
105. L. signifera (Jennings, 1896)
106. L. simonneae Segers, 1993 #
107. L. stenroosi (Meissner, 1908)
108. L. superaculeata Sanoamuang & Segers, 1997
109. L. thienemanni (Hauer, 1938)
110. L. undulata Hauer, 1938
111. L unguitata (Fadeev, 1925)
112. L. ungulata (Gosse, 1887)
Family: Notommatidae
113. Cephalodella gibba (Ehrenberg, 1830)
114. C. mucronata Myers, 1924
115. C. trigona (Rousselet, 1895) *
116. Monommata. grandis Tessin, 1890
117. M. longiseta (O. F. Müller, 1786)
118. Notommata glyphura Wulfert, 1935
119. N. spinata Koste & Shiel, 1991
Family: Scaridiidae
120. Scaridium longicaudum (O. F. Müller, 1786)
Family: Gastropodidae
121. Ascomorpha saltans Bartsch, 1870
Family: Trichocercidae
122. Trichocerca bicristata (Gosse, 1887)
123. T. bidens (Lucks, 1912)
124. T. capucina (Wierzejski & Zacharias, 1893)
125. T. cylindrica (Imhof, 1891)
126. T. elongata (Gosse, 1886)
127. T. flagellata Hauer, 1937
128. T. hollaerti De Smet, 1990 ***
129. T insignis (Herrick, 1885)
130. T. longiseta (Schrank, 1802)
131. T. maior Hauer, 1936 ***
132. T. pusilla (Jennings, 1903)
133. T. rattus (O. F. Müller, 1776)
134. T. scipio (Gosse, 1886)
135. T. similis (Wierzejski, 1893)
136. T. tigris (O. F. Müller, 1786)
Family: Asplanchnidae
137. Asplanchna brightwelli Gosse, 1850
138. A. priodonta Gosse, 1850
Family: Synchaetidae
139. Polyarthra vulgaris Carlin, 1943
Family: Dicranophoridae
140. Dicranophorus forcipatus (O. F. Müller, 1786)
Order: Flosculariaceae
Family: Floscularidae
141. Lacinularia flosculosa (O. F. Müller, 1773)
142. Sinantherina socialis (Linne, 1758)
143. S. spinosa (Thorpe, 1893)
Family: Conochilidae
144. Conochilus unicornis Rousselet, 1892
Family: Hexarthridae
145. Hexarthra mira (Hudson, 1871)
Family: Testudinellidae
146. Testudinella amphora Hauer, 1938 #
147. T. brevicaudata Yamamoto, 1951 #
148. T. dendradena de Beauchamp, 1955 #
149. T. emarginula (Stenroos, 1898)
150. T. greeni Koste, 1981 #
151. T. parva (Ternetz, 1892)
152. T. patina (Hermann, 1783)
153. T. tridentata Smirnov, 1931
154. Pompholyx sulcata Hudson, 1885
Family: Trochosphaeridae
155. Filinia camasecla Myers, 1938 #
156. F. longiseta (Ehrenberg, 1834)
157. F. opoliensis (Zacharias, 1898)
158. F. saltator (Gosse, 1886)
Sub-class: Digononta
Order: Bdelloidea
Family: Philodinidae
159. Dissotrocha aculeata (Ehrenberg, 1832) ***
160. Rotaria neptunia (Ehrenberg, 1830)
* New record from Oriental region; ** New record from northeast India; *** New record from Assam; # reported by Sharma and Khan (2016).
| Arora J, Mehra N K, 2003. Species diversity of planktonic and epiphytic rotifers in the backwaters of the Delhi segment of the Yamuna River, with remarks on new records from India. Zoological Studies, 42(2): 239–247. |
| Bonecker C C, Lansac-Tôha F A, Rossa D C, 1998. Planktonic and non-planktonic rotifers in two environments of the upper Parana River floodplain, state of Mate Grosso do Sul, Brazil. Brazilian Archives of Biology and Technology, 41(4): 447–456. Doi: 10.1590/S1516-89131998000400009 |
| de Paggi S J, 1993. Composition and seasonality of planktonic rotifers in limnetic and littoral regions of a floodplain lake (Paraná River System). Revue d'Hydrobiologie Tropicale, 26(1): 53–63. |
| de Paggi S J, 2001. Diversity of rotifera (Monogononta) in wetlands of Río Pilcomayo national park, Ramsar site(Formosa, Argentina). Hydrobiologia, 462(1-3): 25–34. |
| Dussart B H, Fernando C H, Matsumura-Tundisi J, Shiel R J, 1984. A review of systematics, distribution and ecology of tropical freshwater zooplankton. Hydrobiologia, 113(1): 77–91. Doi: 10.1007/BF00026594 |
| Fernando C H, 1980. The freshwater zooplankton of Sri Lanka, with a discussion of tropical freshwater zooplankton composition. International Review of Hydrobiology, 65(1): 85–125. Doi: 10.1002/(ISSN)1522-2632 |
| Green J, 1972. Latitudinal variation in associations of planktonic Rotifera. Journal of Zoology, 167(1): 31–39. |
| Green J, 2003. Associations of planktonic and periphytic rotifers in a tropical swamp, the Okavango Delta, Southern Africa. Hydrobiologia, 490(1-3): 197–209. |
| Greenberg A E. 1992. Standard Methods: for the Examination of Water and Wastewater. 18th edn. American Public Health Association, Washington D C, USA. |
| Koste W. 1978. Rotatoria. Die rädertiere mitteleuropas. Ein bestimmungswerk, begründet von Max Voigt. Überordnung monogononta. Gebrüder Borntraeger, Berlin, Stuttgart. I. Text (673p. ) U. Ⅱ. Tafelbd. (T. 234). |
| Sanoamuang L O, 1998. Rotifera of some freshwater habitats in the floodplain of the River Nan, northern Thailand. Hydrobiologia, 387-388: 27–33. |
| Segers H, Dumont H J, 1995. 102+ rotifer species (Rotifera:Monogononta) in Broa reservoir (SP., Brazil) on 26 August 1994, with the description of three new species. Hydrobiologia, 316(3): 183–197. Doi: 10.1007/BF00017436 |
| Segers H, Ferrufino N L, de Meester L, 1998. Diversity and Zoogeography of Rotifera (Monogononta) in a flood plain lake of the Ichilo River, Bolivia, with notes on littleknown species. International Review of Hydrobiology, 83(5-6): 439–448. Doi: 10.1002/(ISSN)1522-2632 |
| Segers H, Nwadiaro C S, Dumont H J, 1993. Rotifera of some lakes in the floodplain of the river Niger (Imo State, Nigeria). Ⅱ. Faunal composition and diversity. Hydrobiologia, 250(1): 63–71. Doi: 10.1007/BF00007495 |
| Segers H, Pholpunthin P, 1997. New and rare Rotifera from Thale-Noi Lake, Pattalang Province, Thailand, with a note on the taxonomy of Cephalodella (Notommatidae). Annals of Limnology, 33(1): 13–21. Doi: 10.1051/limn/1997001 |
| Segers H. 1995. Rotifera 2. The Lecanidae. In: Dumont H J, Nogrady T eds. Guides to the Identification of the Microinvertebrates of the Continental Waters of the World 6. SPB Academic Publishing bv. Amsterdam, The Netherlands. p. 1-226. |
| Segers H, 1996. The biogeography of littoral Lecane Rotifera. Hydrobiologia, 323(3): 169–197. Doi: 10.1007/BF00007845 |
| Segers H, 2001. Zoogeography of the Southeast Asian Rotifera. Hydrobiologia, 446-447: 233–246. Doi: 10.1023/A:1017524011783 |
| Segers H, 2007. Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution. Zootaxa, 1564: 1–104. |
| Sharma B K, Dudani V K, 1992. Rotifers from some tropical ponds in Bihar, species composition, similarities and trophic indicators. Journal of the Indian Institute of Science, 72(2): 121–130. |
| Sharma B K, Haokip T P, Sharma S, 2016. Loktak Lake, Manipur, northeast India:a Ramsar site with rich rotifer(Rotifera:Eurotatoria) diversity and its meta-analysis. International Journal of Aquatic Biology, 4(2): 69–79. |
| Sharma B K, Khan S I, 2016. Interesting rotifers (Rotifera:Eurotatoria) from floodplain lakes of lower Brahmaputra river basin of Assam, northeast India. Opuscula Zoologica, Budapest, 47(2): 123–130. |
| Sharma B K, Lyngskor C, 2003. Plankton communities of a subtropical reservoir of Meghalaya (NE India). Indian Journal of Animal Sciences, 73(2): 209–215. |
| Sharma B K, Sharma S, Hatimuria M K, 2015. Rotifer assemblages (Rotifera:Eurotatoria) of the floodplain lakes of Majuli River Island, the Brahmaputra river basin, northeast India. International Journal of Aquatic Biology, 3(1): 1–13. |
| Sharma B K, Sharma S. 1999. Freshwater rotifers (Rotifera: Eurotatoria). In: Fauna of Meghalaya, State Fauna Series, 4(9): 11-161. Zoological Survey of India, Calcutta. |
| Sharma B K, Sharma S, 2001. Biodiversity of Rotifera in some tropical floodplain lakes of the Brahmaputra river basin, Assam (N.E. India). Hydrobiologia, 446-447(1): 305–313. |
| Sharma B K, Sharma S, 2012. Rotifera diversity of a floodplain lake of the Brahmaputra river basin of lower Assam, Northeast India. Opuscula Zoologica, Budapest, 43(1): 67–77. |
| Sharma B K, Sharma S. 2014a. Northeast India-an important region with a rich biodiversity of Rotifera. In: Sharma B K, Dumont H J, Wallace R L eds. Rotifera XⅢ: Rotifer Biology-A Structural and Functional Approach. International Review of Hydrobiology, 99(1-2): 20-37. |
| Sharma B K, Sharma S. 2014b. Floodplains of the Brahmaputra river basin-globally interesting ecotones with rich Rotifer(Rotifera: Eurotatoria) biodiversity. In: Sinha R K, Ahmed B eds. Rivers for Life-Proceedings of the International Symposium on River Biodiversity: Ganges-BrahmaputraMeghan River System, Ecosystems for Life, A Bangladesh-India Initiative, IUCN, p. 258-270. |
| Sharma B K, Sharma S, 2014c. Indian Lecanidae (Rotifera:Eurotatoria:Monogononta) and its distribution. In:Sharma B K, Dumont H J, Wallace R L eds. Rotifera Xó:Rotifer Biology-a structural and functional Approach. International Review of Hydrobiology, 99(1-2): 38–47. |
| Sharma B K, Sharma S, 2015a. New records of rotifers(Rotifera:Eurotatoria) from Deepor beel-a Ramsar site of India with an update on its rich Rotifer diversity. Journal of Threatened Taxa, 7(3): 7. |
| Sharma B K, Sharma S, 2015b. Biodiversity of freshwater rotifers (Rotifera:Eurotatoria) of Mizoram, Northeast India:composition, new records and interesting features. International Journal of Aquatic Biology, 3(5): 301–313. |
| Sharma B K, 1979. Further contributions to the lecanid fauna(Rotifera:Lecanidae) of West Bengal. Acta Hydrobiologica, 21(1): 53–59. |
| Sharma B K. 1998. Freshwater rotifers (Rotifera: Eurotatoria). In: Fauna of West Bengal. State Fauna Series, 3(11): 341-461. Zoological Survey of India, Calcutta, India. |
| Sharma B K, 2005. Rotifer communities of floodplain lakes of the Brahmaputra basin of lower Assam (N.E. India):biodiversity, distribution and ecology. Hydrobiologia, 533(1): 209–221. |
| Sharma B K, 2009a. Diversity of rotifers (Rotifera, Eurotatoria) of Loktak lake, Manipur, North-Eastern India. Tropical Ecology, 50(2): 277–285. |
| Sharma B K, 2009b. Rotifer communities of floodplain lakes of Manipur (North-East India):biodiversity, distribution and ecology. Journal of the Bombay Natural History Society, 106(1): 45–56. |
| Sharma B K, 2011. Zooplankton diversity of two floodplain lakes (pats) of Manipur, northeast India. Opuscula Zoologica Budapest, 42(2): 185–197. |
| Sharma B K, 2014. Rotifers (Rotifera:Eurotatoria) from wetlands of Majuli-the largest river island, the Brahmaputra river basin of upper Assam, northeast India. Check List, 10(2): 292–298. Doi: 10.15560/10.2.292 |
| Sharma B K, 2016. Interesting rotifers (Rotifera:Eurotatoria) from a subtropical wetland of Meghalaya, Northeast India:new records. Turkish Journal of Zoology, 40(3): 433–437. |
| Sharma S, Sharma B K, 2008. Zooplankton diversity in floodplain lakes of Assam. Records of the Zoological Survey of India, Occasional Paper No., 290: 1–307. |
| Sharma S, Sharma B K, 2013. Faunal diversity of aquatic invertebrates of Deepor Beel (a Ramsar site), Assam, northeast India. Wetland Ecosystem Series, 17: 1–226. |
| Zoological Survey of India, Kolkata. Shiel R J, Green J D, Nielsen D L, 1998. Floodplain biodiversity:why are there so many species?. Hydrobiologia, 387-388: 39–46. |
| Sládeček V, 1983. Rotifers as indicators of water quality. Hydrobiologia, 100(1): 169–201. Doi: 10.1007/BF00027429 |
| Sugunan V V. 1997. Fisheries management of small water bodies in seven countries in Africa, Asia and Latin America. Food & Agriculture Organization Circular No. 933. FAO, Rome. p. 1-149. |