 2019, Vol. 37
2019, Vol. 37Institute of Oceanology, Chinese Academy of Sciences
Article Information
- SUN Tingting, DONG Zhijun, LI Yuanchao
- Versuriga anadyomene, a newly recorded scyphozoan jellyfish (Scyphozoa: Rhizostomae) in Chinese waters
- Journal of Oceanology and Limnology, 37(1): 266-272
- http://dx.doi.org/10.1007/s00343-018-7273-8
Article History
- Received Oct. 22, 2017
- accepted in principle Dec. 11, 2017
- accepted for publication Feb. 22, 2019




2 Hainan Academy of Ocean and Fisheries Sciences, Haikou 570203, China
The class Scyphozoa includes approximately 201 accepted species worldwide (Mayer, 1910; Kramp, 1961; Daly et al., 2007; Appeltans et al., 2012; Daglio and Dawson, 2017). Only 35 species of scyphomedusae, belonging to 20 genera in 16 families, have been recorded in Chinese seas. Of these, 7 species belong to the order Coronatae, 8 to Semaeostomeae and 20 to Rhizostomeae (Gao et al., 2002; Hong and Lin, 2010). Recently, these scyphozoans have caused great concern due to the effect of their blooms on fisheries, aquaculture, tourism, and coastal plants (Purcell, 2012). Three scyphozoan species, Aurelia aurita, Cyanea nozakii and Nemopilema nomurai, form large blooms in Chinese seas (Dong et al., 2010).
Recent research indicates that scyphozoans are preyed upon by other organisms, such as leatherback sea turtles, penguins, bluefin tuna, and thus could play an important role in the marine food web (Cardona et al., 2015; Hamilton, 2016; Marques et al., 2016; Bos et al., 2017). Furthermore, scyphozoans are considered fisheries resources and can be utilized as a food source in Asia (Hsieh, 2001; Kitamura and Omori, 2010; Brotz et al., 2017). For example, approximately 0.19–0.33 million tonnes of the edible jellyfish Rhopilema esculentum per year were caught for food in China between 2000 and 2011 (Dong et al., 2014).
During a field survey in the South China Sea, samples of rhizostome jellyfish which had not been previously recorded in Chinese seas were collected. These specimens were identified as Versuriga anadyomene based on a combination of morphological characteristics and 18S ribosomal DNA sequences data. This is the first record of Versuriga anadyomene in Chinese coastal waters.
2 MATERIAL AND METHODThree rhizostome jellyfish specimens were collected with a hand net in Beibu Gulf, South China Sea on 27 May 2015. The underwater photographs of the medusae were taken using a Canon PowerShot G1X digital camera with an underwater housing. The morphological characters were examined according to the previous studies (Mayer, 1910; Kramp, 1961; Gershwin et al., 2010). In addition, medusa tissue, clipped from the bell margin or gonads, was preserved in 95% ethanol and stored at -20℃ until DNA extraction.
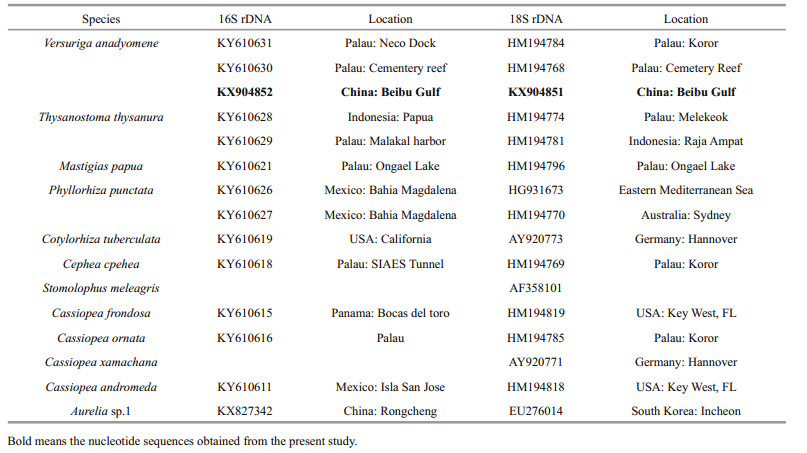
Total genomic DNA was extracted using the TIANamp Marine Animals DNA Kit (TIANGEN, China). The mitochondrial cytochrome c oxidase subunit Ⅰ (COI) fragments were amplified using the universal primers LCO1490 (GGTCAACAAATCA-TAAAGATATTGG) and HCO2198 (TAAACTTCA-GGGTGACCAAAAAATCA) under the PCR conditions previously described (Folmer et al., 1994). The mitochondrial 16S rDNA (16S) fragments were amplified using the primers 16S-L (GACTGTTTA-CCAAAAACATA) and 16S-H (CATAATTCAACA-TCGAGG) under the PCR conditions previously described (Ender and Schierwater, 2003). The nuclear 18S rDNA fragments were amplified using the primers 18S-L (5′-AACCTGGTTGATCCTGCCA-GT-3′) and 18S-H (5′-GATCCTTCTGCAGGTTCA-CCTAC-3′) under the PCR conditions described by Medlin et al. (1988) and Collins (2002).
The PCR products were examined on 1% agarose gels, stained with GenecolourTM (Biotium, USA), and photographed with transmitted illumination. The PCR-amplified DNA fragments were purified using a DNA Gel Extraction Kit (Sangon, China). The purified PCR products were cloned using a pUCm-T vector kit (Sangon Biotech Co., Shanghai, China), and then sequenced with an ABI 3730 automated DNA sequencer at Sangon Biotech Co. (Shanghai, China). A BLAST search for the sequences was performed in the NCBI database to confirm their identities. In addition, related mitochondrial 16S rDNA and nuclear 18S rDNA gene sequences for Scyphozoa were downloaded from NCBI and aligned with the new sequence (Table 1). The mitochondrial 16S rDNA and nuclear 18S rDNA gene sequence of Aurelia sp.1 (KX827342 for 16S rDNA, EU276014 for 18S rDNA) was used as an outgroup. Sequences were aligned with the program Clustal X. Maximum likelihood analysis of mitochondrial 16S rDNA and nuclear 18S rDNA data was performed based on the GTR+G+I model and TN93+G+I model, respectively, which were selected using the Akaike information criterion (Akaike, 1992). Bootstrap values were calculated using 1000 iterations. All phylogenetic analyses were performed using MEGA 5.0 software (Tamura et al., 2011).
|
Class SCYPHOZOA Goette, 1887
Order RHIZOSTOMEAE Cuvier, 1800
Family VERSURIGIDAE Kramp, 1961
Genus Versuriga Kramp, 1961
Versuriga anadyomene (Maas, 1903)
3.2 Morphological charactersFigure 1 shows the morphological characters of the specimens we collected. These specimens had a hemispherical umbrella, up to 600 mm in diameter, with reticular grooves on the exumbrella surface. The margin of the umbrella bore eight large semicircular velar lappets, alternating with narrow rhopaliar lappets in each of the 8 octants. The mouth-arms were flattened laterally, with numerous flat, membranous branches bearing many clubs and intermediate filaments. Subumbrella canal structures included non-anastomosed broad perradial canals and anastomosed narrow interradial canals.

|
| Fig.1 Versuriga anadyomene from Beibu Gulf, South China Sea Bell diameter is approximately 550 mm. a. lateral view; b. reticular grooves on the exumbrella surface; c. oral view, showing semicircular velar lappets; d. narrow rhopaliar lappets; e. membranous branches bearing many clubs and intermediate filaments; f. oral view, showing subumbrella canal. |
The partial sequences of mitochondrial COI, mitochondrial 16S rDNA, and nuclear 18S rDNA genes were deposited in GenBank (GenBank accession numbers KX904853 for COI, KX904852 for 16S rDNA, and KX904851 for 18S rDNA). The best match for the mitochondrial COI gene sequence in the NCBI is only 85% identical. A BLAST search of the GenBank database revealed that the mitochondrial 16S rDNA sequences from Beibu Gulf were 99% identical to the 16S rDNA sequences of V. anadyomene from Palau (KY610630, KY6106301) (Daglio and Dawson, 2017). Phylogenetic analysis of aligned 16S rDNA gene sequences indicated that our sequence from Beibu Gulf clustered in the same group as V. anadyomene collected from Palau; this was supported by a 96% bootstrap value in ML trees (Fig. 2). Additionally, the 18S rDNA sequences from Beibu Gulf were 99% identical to the 18S rDNA sequences of V. anadyomene from Palau (HM194768, HM194784) (Bayha et al., 2010). Phylogenetic analysis of aligned 18S rDNA gene sequences indicated that our sequence clustered in the same group as V. anadyomene collected from Palau; this was supported by a 98% bootstrap value in ML trees (Fig. 3).

|
| Fig.2 Maximum-likelihood tree constructed using GTR+G+I model for mitochondrial 16S rDNA gene sequences of V. anadyomene and other scyphozoans |

|
| Fig.3 Maximum-likelihood tree constructed using TN93+G+I model for nuclear 18S rDNA gene sequences of V. anadyomene and other scyphozoans |
The rhizostome jellyfish V. anadyomene belongs to phylum Cnidaria, class Scyphozoa, order Rhizostomeae, family Versurigidae, genus Versuriga (Kramp, 1961). The adult medusae of V. anadyomene are large, reaching up to 60 cm across the umbrella. This species has been reported in the coastal waters of Malaysia (Maas, 1903), the Philippines (Mayer, 1910), Indonesia (Ohtsuka et al., 2009), Palau (Bayha et al., 2010), and Australia (Gershwin et al., 2010). The morphological characters of the rhizostome jellyfish collected in the Beibu Gulf, South China Sea, were consistent with published descriptions of V. anadyomene (Mayer, 1910; Kramp, 1961; Gershwin et al., 2010). This was further confirmed by the phylogenetic analysis of mitochondrial 16S rDNA and nuclear 18S rDNA gene fragments. This study represents the first record of the species Versuriga anadyomene in Chinese seas.
Most scyphozoans have a metagenic life cycle, alternating between sexually reproducing pelagic medusae and sessile asexual polyps (Arai, 2001). The dispersal of the scyphozoans can be achieved in two ways: local dispersal by physical transport in the pelagic medusa stage and global transport via ships, barges, and offshore drilling platforms in the benthic polyp stage (Dawson et al., 2005; Bayha and Graham, 2009). Recently, a growing number of newly-recorded scyphozoan species have been reported in different areas of the world's oceans (e.g., Çevik et al., 2006; Gershwin and Zeidler, 2008; Ocaña-Luna et al., 2010; Yahia et al., 2013; Deidun et al., 2017; Morandini et al., 2017). Some species were confirmed as invasive or alien species, including Phyllorhiza punctata, Rhopilema nomadica, Marivagia stellata, Cotylorhiza erythraea, and Cassiopea andromeda in the Mediterranean (Çevik et al., 2006; Galil et al., 2009, 2010; Yahia et al., 2013; Deidun et al., 2017), and P. punctata in the Gulf of Mexico (Ocaña-Luna et al., 2010). Dispersal in the South China Sea is not restricted, and thus we infer that the rhizostome jellyfish V. anadyomene in the Beibu Gulf may have been transported from the waters of Southeast Asia, i.e., Malaysia, the Philippines, or Indonesia. It is unknown if V. anadyomene has established populations in the Beibu Gulf, due to the absence of information on the polyp and ephyra stages of this species.
Jellyfish populations are increasing in many areas of the world and are considered as nuisance to fisheries, aquaculture, coastal power plants, and tourism (Dong et al., 2010; Purcell, 2012). However, some species of scyphozoan jellyfish are also edible and have been exploited commercially as an important food (Hsieh et al., 2001). As many as 35 jellyfish species have been reported to be consumed by humans, most of which are belonging to the Order Rhizostomeae (Brotz et al., 2017). The rhizostome jellyfish V. anadyomene has tough and rigid tissue and thus can be utilized to produce processed food products. Future studies on the population dynamics and stock resources are needed to assess the possibility of exploiting edible products from the jellyfish.
5 CONCLUSIONThe genus Versuriga and V. anadyomene has been recorded for the first time in Chinese coastal waters based on morphological characters and DNA sequences data. The morphological characters of these rhizostome jellyfish were consistent with descriptions of specimens from other areas of Southeast Asia. Phylogenetic analysis of mitochondrial 16S rDNA and nuclear 18S rDNA gene fragments confirmed that these specimens were in the same cluster as V. anadyomene collected in Palau. Future studies on the population dynamics and stock resources are needed to assess the possibility of exploiting edible products from the jellyfish.
Akaike H. 1992. Information theory as an extension of the maximum likelihood principle. In: Proceedings of the 2nd International Symposium on Information Theory.Academiai, Budapest, Hungaty. p.267-281.
|
Appeltans W, Ahyong S T, Anderson G, Angel M V, Artois T, Bailly N, Bamber R, Barber A, Bartsch I, Berta A, Błażewicz-Paszkowycz M, Bock P, Boxshall G, Boyko C B, Brandão S N, Bray R A, Bruce N L, Cairns S D, Chan T Y, Cheng L N, Collins A G, Cribb T, Curini-Galletti M, Dahdouh-Guebas F, Davie P J F, Dawson M N, De Clerck O, Decock W, De Grave S, De Voogd N J, Domning D P, Emig C C, Erséus C, Eschmeyer W, Fauchald K, Fautin D G, Feist S W, Fransen C H J M, Furuya H, GarciaAlvarez O, Gerken S, Gibson D, Gittenberger A, Gofas S, Gómez-Daglio L, Gordon D P, Guiry M D, Hernandez F, Hoeksema B W, Hopcroft R R, Jaume D, Kirk P, Koedam N, Koenemann S, Kolb J B, Kristensen R M, Kroh A, Lambert G, Lazarus D B, Lemaitre R, Longshaw M, Lowry J, Macpherson E, Madin L P, Mah C, Mapstone G, McLaughlin P A, Mees J, Meland K, Messing C G, Mills C E, Molodtsova T N, Mooi R, Neuhaus B, Ng P K L, Nielsen C, Norenburg J, Opresko D M, Osawa M, Paulay G, Perrin W, Pilger J F, Poore G C B, Pugh P, Read G B, Reimer J D, Rius M, Rocha R M, Saiz-Salinas J I, Scarabino V, Schierwater B, Schmidt-Rhaesa A, Schnabel K E, Schotte M, Schuchert P, Schwabe E, Segers H, SelfSullivan C, Shenkar N, Siegel V, Sterrer W, Stöhr S, Swalla B, Tasker M L, Thuesen E V, Timm T, Todaro M A, Turon X, Tyler S, Uetz P, Van Der Land J, Vanhoorne B, Van Ofwegen L P, Van Soest R W M, Vanaverbeke J, Walker-Smith G, Walter T C, Warren A, Williams G C, Wilson S P, Costello M J. 2012. The magnitude of global marine species diversity. Curr. Biol., 22(23): 2 189-2 202.
DOI:10.1016/j.cub.2012.09.036 |
Arai M N. 2001. Pelagic coelenterates and eutrophication: a review. In: Purcell J E, Graham W M, Dumont H J eds.Jellyfish Blooms: Ecological and Societal Importance.Springer, Dordrecht, Netherlands. p.69-87.
|
Bayha K M, Dawson M N, Collins A G, Barbeitos M S, Haddock S H D. 2010. Evolutionary relationships among scyphozoan jellyfish families based on complete taxon sampling and phylogenetic analyses of 18S and 28S ribosomal DNA. Integr. Comp. Biol., 50(3): 436-455.
DOI:10.1093/icb/icq074 |
Bayha K M, Graham W M. 2009. A new Taqman© PCR-based method for the detection and identification of scyphozoan jellyfish polyps. Hydrobiologia, 616(1): 217-228.
DOI:10.1007/s10750-008-9590-y |
Bos A R, Cruz-Rivera E, Sanad A M. 2017. Herbivorous fishes Siganus rivulatus (Siganidae) and Zebrasoma desjardinii(Acanthuridae) feed on ctenophora and scyphozoa in the Red Sea. Mar. Biodiver., 47(1): 243-246.
|
Brotz L, Schiariti A, López-Martínez J, Álvarez-Tello J, Hsieh Y H P, Jones R P, Quiñones J, Dong Z J, Morandini A C, Preciado M, Laaz E, Mianzan H. 2017. Jellyfish fisheries in the Americas:origin, state of the art, and perspectives on new fishing grounds. Rev. Fish Biol. Fish, 27(1): 1-29.
DOI:10.1007/s11160-016-9445-y |
Cardona L, Martínez-Iñigo L, Mateo R, González-Solís J. 2015. The role of sardine as prey for pelagic predators in the western Mediterranean Sea assessed using stable isotopes and fatty acids. Mar. Ecol. Prog. Ser., 531: 1-14.
DOI:10.3354/meps11353 |
Çevik C, Erkol I L, Toklu B. 2006. A new record of an alien jellyfish from the levantine coast of turkey-Cassiopea andromeda (Forsskål, 1775)[Cnidaria:Scyphozoa:Rhizostomea]. Aquat. Invasions, 1(3): 196-197.
|
Collins A G. 2002. Phylogeny of Medusozoa and the evolution of cnidarian life cycles. J. Evol. Biol., 15(3): 418-432.
DOI:10.1046/j.1420-9101.2002.00403.x |
Daglio L G, Dawson M N. 2017. Species richness of jellyfishes(Scyphozoa:Discomedusae) in the Tropical Eastern Pacific:missed taxa, molecules, and morphology match in a biodiversity hotspot. Invertebr. Syst., 31(5): 635-663.
DOI:10.1071/IS16055 |
Daly M, Brugler M R, Cartwright P, Collins A G, Dawson M N, Fautin D G, France S C, McFadden C S, Opresko D M, Rodriguez E, Romano S L, Stake J L. 2007. The phylum Cnidaria:a review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa, 1668: 127-182.
|
Dawson M N, Gupta A S, England M H. 2005. Coupled biophysical global ocean model and molecular genetic analyses identify multiple introductions of cryptogenic species. Proc. Natl. Acad. Sci. USA, 102(34): 11 968-11 973.
DOI:10.1073/pnas.0503811102 |
Deidun A, Sciberras J, Sciberras A, Gauci A, Balistreri P, Salvatore A, Piraino S. 2017. The first record of the whitespotted Australian jellyfish Phyllorhiza punctata von Lendenfeld, 1884 from Maltese waters (western Mediterranean) and from the Ionian coast of Italy. BioInvasions Rec., 6(2): 119-124.
|
Dong Z J, Liu D Y, Keesing J K. 2010. Jellyfish blooms in China:dominant species, causes and consequences. Mar.Pollut. Bull., 60(7): 954-963.
DOI:10.1016/j.marpolbul.2010.04.022 |
Dong Z J, Liu D Y, Keesing J K. 2014. Contrasting trends in populations of Rhopilema esculentum and Aurelia aurita in Chinese waters. In: Pitt K A, Lucas C H eds. Jellyfish Blooms. Springer, Dordrecht, Netherlands. p.207-218.
|
Ender A, Schierwater B. 2003. Placozoa are not derived cnidarians:evidence from molecular morphology. Mol.Biol. Evol., 20(1): 130-134.
DOI:10.1093/molbev/msg018 |
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol., 3(5): 294-299.
|
Galil B S, Gershwin L A, Douek J, Rinkevich B. 2010. Marivagia stellata gen. et sp. nov. (Scyphozoa:Rhizostomeae:Cepheidae), another alien jellyfish from the Mediterranean coast of Israel. Aquat. Invasions, 5(4): 331-340.
|
Galil B S, Shoval L, Goren M. 2009. Phyllorhiza punctata von Lendenfeld, 1884 (Scyphozoa:Rhizostomeae:Mastigiidae) reappeared off the Mediterranean coast of Israel. Aquat. Invasions, 4(3): 481-483.
|
Gao S W, Hong H X, Zhang S M. 2002. Fauna Sinica, Invertebrata Vol. 27. Phylum Cnidaria, Class Hydrozoa, Subclass Siphonophorae. Class Scyphomedusae. Science Press, Beijing, China. (in Chinese)
|
Gershwin L A, Zeidler W, Davie P J F. 2010. Medusae(Cnidaria) of Moreton Bay, Queensland, Australia. Mem.Qld. Mus, 54(3): 47-108.
|
Gershwin L A, Zeidler W. 2008. Some new and previously unrecorded Scyphomedusae (Cnidaria:Scyphozoa) from southern Australian coastal waters. Zootaxa, 1744: 1-18.
DOI:10.11646/zootaxa.1744.1 |
Hamilton G. 2016. The secret lives of jellyfish:long regarded as minor players in ocean ecology, jellyfish are actually important parts of the marine food web. Nature, 531(7595): 432-434.
DOI:10.1038/531432a |
Hong H X, Lin L M. 2010. Study on the fauna of the scyphomedusae in the China Sea. J. Jimei Univ. (Nat.Sci.), 15(1): 18-24.
(in Chinese with English abstract) |
Hsieh Y P H, Leong F M, Rudloe J. 2001. Jellyfish as food. Hydrobiologia, 451(1-3): 11-17.
|
Kitamura M, Omori M. 2010. Synopsis of edible jellyfishes collected from Southeast Asia, with notes on jellyfish fisheries. Plankton Benthos Res., 5(3): 106-118.
DOI:10.3800/pbr.5.106 |
Kramp P L. 1961. Synopsis of the medusae of the world. J.Mar. Biol. Assoc. UK, 40: 7-382.
DOI:10.1017/S0025315400007347 |
Marques R, Bouvier C, Darnaude A M, Molinero J C, Przybyla C, Soriano S, Tomasini J A, Bonnet D. 2016. Jellyfish as an alternative source of food for opportunistic fishes. J.Exp. Mar. Biol. Ecol., 485: 1-7.
DOI:10.1016/j.jembe.2016.08.008 |
Mass O. 1903. Die Scyphomedusen der Siboga-expedition. Leiden, Buchhandlung und Druckerei Vormals E. J. Brill.Siboga Exped., 11(9): 1-91.
|
Mayer A G. 1910. Medusae of the World. Vol Ⅲ: Scyphozoa.Carnegie Institute, Washington, USA.
|
Medlin L, Elwood H J, Stickel S, Sogin M L. 1988. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene, 71(2): 491-499.
DOI:10.1016/0378-1119(88)90066-2 |
Morandini A C, Stampar S N, Maronna M M, Da Silveira F L. 2017. All non-indigenous species were introduced recently? The case study of Cassiopea (Cnidaria:Scyphozoa) in Brazilian waters. J. Mar. Biol. Assoc. UK, 97(2): 321-328.
|
Ocaña-Luna A, Sánchez-Ramírez M, Aguilar-Durán R. 2010. First record of Phyllorhiza punctata von Lendenfeld, 1884 (Cnidaria:Scyphozoa, Mastigiidae) in Mexico. Aquat. Invasions, 5(S1): S79-S84.
|
Ohtsuka S, Koike K, Lindsay D, Nishikawa J, Miyake H, Kawahara M, Mulyadi, Mujiono N, Hiromi J, Komatsu H. 2009. Symbionts of marine medusae and ctenophores. Plankton Benthos Res., 4(1): 1-13.
DOI:10.3800/pbr.4.1 |
Purcell J E. 2012. Jellyfish and ctenophore blooms coincide with human proliferations and environmental perturbations. Ann. Rev. Mar. Sci., 4: 209-235.
DOI:10.1146/annurev-marine-120709-142751 |
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol.Evol., 28(10): 2 731-2 739.
DOI:10.1093/molbev/msr121 |
Yahia M N D, Yahia O K D, Gueroun S K M, Aissi M, Deidun A, Fuentes V, Piraino S. 2013. The invasive tropical scyphozoan Rhopilema nomadica Galil, 1990 reaches the Tunisian coast of the Mediterranean Sea. BioInvasions Rec., 2(4): 319-323.
|