 2019, Vol. 37
2019, Vol. 37Institute of Oceanology, Chinese Academy of Sciences
Article Information
- MA Qin, LI Mingzheng, QIN Xiangchao, LIU Huanzhang, CAO Wenxuan
- Ichthyoplankton recruitment from mainstream of the Changjiang River into the Dongting Lake, the second largest freshwater lake in China
- Journal of Oceanology and Limnology, 37(3): 1090-1101
- http://dx.doi.org/10.1007/s00343-019-8036-x
Article History
- Received Mar. 2, 2018
- accepted in principle May. 21, 2018
- accepted for publication Aug. 3, 2018




2 University of Chinese Academy of Sciences, Beijing 100049, China;
3 Yellow River Basin Water Environment Monitoring Center, Zhengzhou 450004, China
Riverine floodplains are among the Earth's most distinctive landscapes, characterized by their high biodiversity and productivity and their corresponding recreational and aesthetic values (Tockner and Stanford, 2002). Floodplains are also considered centers of biocomplexity and bioproduction, with more species of plants and animals occurring on floodplains than in any other landscape unit in most regions of the world (Tockner and Stanford, 2002). As a typical example, the Changjiang (Yangtze) River and its floodplain lakes were listed as one of the seven large river ecosystems with the highest biodiversity and endemicity in the world (Martens and Segers, 2009) and suggested as hotspots of freshwater biodiversity (Kottelat and Whitten, 1996) and one of the 200 global conservation priority ecoregions by the World Wide Fund (Olson and Dinerstein, 2002).
A key feature of the Changjiang floodplain lakes is their high diversity of fish species. More than a thousand lakes occupy the middle Changjiang floodplain, and almost all of these lakes are rich in fish species. For example, 134 species are found in Poyang Lake (Fang et al., 2016), 116 species in Dongting Lake (Li et al., 2013a), and 87 species in Wuhu Lake (Liang et al., 1981). This high diversity is attributed to the river-lake migratory species (RL). After a long geological period, as adaptation to the East Asian monsoon climate, the Asian cypriniforms, such as the Chinese major carps, evolved spawning of drift eggs (Cao, 2011). When water temperature is above 18℃ and with flooding resulting in a water rise > 0.55 m/day, these fishes may migrate to the spawning ground and spawn drift eggs (Yi et al., 1988a; Li et al., 2013b). The eggs need to float for a certain duration until they hatch, then the larvae mature to the swimming stage. This development process needs a specific duration and flow velocity; thus, successful spawning can only occur in large rivers (Li et al., 2013b) and never in lakes. Given that lakes can provide rich food, larvae and juveniles of river spawning fishes often enter the affiliated lakes and grow there. Thus, these river-lake migratory fishes significantly increase fish diversity in lakes (Ru et al., 2008).
River-lake migratory fishes are important components of lake ecosystems. For example, in Dongting Lake, river-lake migratory fishes once accounted for 32% of the catch of commercial fisheries (Liao et al., 2002), and in Wuhu Lake, they can reach as high as 77.6% (Liang et al., 1981). Liang et al. (1981) documented ~40 species with river-lake migration or semi-migration behavior. Without these river-lake migratory species, fish diversity in lakes will drop sharply and the fish community will remarkably differ. Potentially, environmental changes such as eutrophication could occur in the lakes (Fang et al., 2006; Liao et al., 2006). Despite the importance of river-lake fishes to the lake ecosystem, the recruitment mechanisms of river-lake fish ichthyoplankton have not been investigated thoroughly.
Dongting Lake (28°44′–29°35′N; 111°53′–113°05′E), located in Central China, is the second largest freshwater lake in China. With approximately 1 428 plant species, 116 fishes, 217 birds, and the unique finless porpoise (Neophocaena asiaeorientalis asiaeorientalis) (Li and Shen, 2005; Li et al., 2013a), this lake is famous for its valuable and exclusive biodiversity. The fisheries catches alone, account for almost 1/4 to 1/3 of the total amount of fishes in the Changjiang River (Chen et al., 2002; Xie, 2017), and therefore bears importance in the fish industry. This lake also serves as an important water supply and helps with flood mitigation, wastewater treatment, and transportation.
Dongting Lake receives water from four tributaries (Xiangjiang, Yuanjiang, Zishui, and Lishui Rivers) and through three inlet channels (Songzi, Ouchi, and Taiping channels) of the Changjiang mainstream, but it only features one outlet channel (Chenglingji) to the Changjiang River (Dou and Jiang, 2000). Historically, ichthyoplankton recruitment is roughly estimated as the percentage of water discharge flowing through the three inlet channels and varies between 5.2% to 11.8% for the different channels (Zhou et al., 1986). However, in situ field investigations have never been conducted. In recent decades, the inlet channels shrank, and siltation occurred due to heavy human activities, such as flow regulation, damming, and agricultural practices or extractions, resulting in lower water discharge flowing into the Dongting Lake (Han and Zhou, 1999; Hu et al., 2014; Sun and Shen, 2015).
In this study, we surveyed the ichthyoplankton flowing into the Dongting Lake through the largest of its three inlet channels, the Songzi channel. We aimed to assess the current situation and temporal variations of ichthyoplankton recruitment to the Dongting Lake and determine the environmental factors related to ichthyoplankton production to provide suggestions for future fish resource conservation and management.
2 MATERIAL AND METHOD 2.1 Study areaCurrently, Songzi channel is the largest floodway channel connecting the Changjiang River mainstream and the Dongting Lake. On average 12% of the mainstream discharge into the Dongting Lake via the Songzi channel, which in turn plays an important role in fish recruitment to the Dongting Lake (Sun and Shen, 2015).
The Songzi channel, with a length of 353 km, is primarily divided into east and west branches. The samples were collected in the west branch of Songzi channel. The sampling site was situated in Songzi city, about 37 km away from the inlet (111°77'E, 30°17'N). The channel is 220–270 m wide, with corresponding discharge of 350–3 300 m3/s from May to July. Sampling was conducted 10–60 m away from the bank, and the depth ranged from 2 m to 8 m depending on water discharge (Fig. 1).

|
| Fig.1 Map of the Changjiang floodplain showing the study area on the middle Changjiang River The black triangle indicates the sampling location. |
Fish eggs and larvae were collected daily from May 23 to July 24 in 2013 and from May 1 to July 26 in 2014 in the Songzi channel. Routine sampling was conducted twice a day, generally from 6:00 to 7:30 and 17:30 to 19:00. Each collection lasted for 15–60 min, depending on water transparency and velocity.
A semi-conical net was used to collect eggs and larvae (Yi et al., 1988a, Duan et al., 2009); it had a 0.39-m2 semi-circular mouth opening, was 2 m in length with a mesh-size of 500 μm and was placed just below the water surface (0.5 m water depth). A collection box (40 cm×30 cm×30 cm) at the end of the net, was used to collect eggs and larvae. The semiconical net and the collection box were made of thin silk like material. The upper part of the box was open to allow removal of samples and remained above the water surface by buoyancy. The sampling net was fixed to a shipboard with ropes and kept perpendicular to the current.
To standardize the results of routine sampling, cross-section sampling (as Mu et al. (2014) defined as special sampling) was conducted along a transect across the river once or twice each sampling period. This method can obtain the distribution coefficient ("C", see formulae in the "Data analysis" section) of fish eggs and larvae in the sampling section and was used to estimate the total number of eggs and larvae in subsequent computations (Yi et al., 1988a; Humphries et al., 2002; Duan et al., 2009; Mu et al., 2014; Li et al., 2016). We used a conical net (2 m long, 0.5 m in diameter, with 500 μm in mesh size) for cross-section sampling (Cao et al., 2007). Five sites were specifically selected evenly along the transect (next to the left bank, halfway between the left bank and the river center, the central midpoint of the river, halfway between the central midpoint and the right bank and next to the right bank). At each site three water depths were sampled (0.2, 0.5, and 0.8 times of the depths under the water level in the sampling site; Mu et al. (2014)). The routine sampling site was next to the right bank.
2.3 Sample processingAfter sampling, fish eggs and larvae were separated and counted immediately, and the stages of development were identified. The stages of development and developmental time of each stage were defined according to the method of Yi et al. (1988b). According to development, the eggs were incubated in paper cups for five to eight days and identified at the species level. Each cup was stocked with one to ten eggs which were at the same development stage. During incubation, water temperature was maintained between 22℃ and 26℃ by air conditioner, and the water was aerated (see also Li et al., 2013b). Species were identified under a stereomicroscope to the lowest practical taxa according to the works of Yi et al. (1988b) and Cao et al. (2007).
2.4 Definition of fishes with different ecotypesBased on ecotype, the collected fishes can be categorized into three types, namely, river-lake migratory species (RL), lake-residence species (LS), and riverine species (RI). River-lake migratory species spawn in rivers but use lakes as their nursery ground due to the rich food resource or spawn in lakes but overwinter in the mainstream. Lake-residence species stay in lakes during their whole life stage with no evident migration. Riverine species usually live in rivers but occasionally move to lakes (Chang and Cao, 1999).
2.5 Environmental data collectionEnvironmental parameters were measured by YSI multi-parameter water quality meter on each sampling occasion, including water temperature (℃), water velocity (m/s), dissolved oxygen, conductivity, water hardness, and oxidation-reduction potential were measured. Water transparency was measured by a secchi disk. The daily water discharge and water level data from the Songzi gauge station (0.5 km downstream from sampling site), were downloaded from the National Water and Rainfall Information Network (http://xxfb.hydroinfo.gov.cn/index.html).
2.6 Data analysisDensities of fish eggs and larvae were calculated as number of organisms/1 000 m3. Water volume was calculated by area of the net entrance (m2), mean current velocity at the net entrance (m/s), and sampling duration (s).
The total number of eggs and larvae were calculated according to the following formulae (see also Mu et al., 2014; Li et al., 2016):
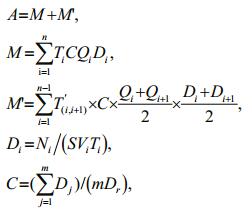
where A refers to the total number of eggs and larvae that drifted through the sampled river section during the entire sampling season; M represents the total number of eggs and larvae counted during all routine samplings; M' stands for the total number of eggs and larvae collected during the time interval between each two adjacent routine sampling occasions; n denotes the number of routine samplings; i corresponds to one specific routine sampling; i+1 indicates the next sampling after the ith sampling; Qi is water discharge of the river reach during the ith sampling (m3/s); Di specifies the drifting density of eggs and larvae during the specific ith sampling (ind./m3); C expresses the distribution coefficient of fish eggs and larvae in the sampled section; T(i, i+1) signifies the time interval between the ith sampling and (i+1)th sampling (s); Ni identifies the number of fish eggs and larvae collected during the ith sampling; S describes net mouth area (m2); Vi depicts water velocity (m/s) at the net mouth during the ith sampling; Ti is duration of the specific ith sampling (s); Dj denotes the density of eggs and larvae at each sampling site as determined by sampling on the transect (ind./m3); j is the specific transect sampling site; m is the number of sampling sites when sampling on the transect; and Dr symbolizes the density of eggs and larvae in the surface of the site next to the right bank when sampling on the transect (ind./m3).
Canonical correspondence analysis (CCA) was used to examine the relationships between the ichthyoplankton drift density and environmental variables. To remove highly correlated variables, all environmental variables whose inflation factors reached more than 20 should be screened. A stepwise process, the manual forward-selection procedure, was used to identify the minimal set of environmental variables that can best explain the variation in species data (Ter Braak, 1988). Forward selection sequentially tested the statistical significance of environmental variables that contributed most strongly to the canonical model. Selected environmental variables were added to the model when their F-ratios were as high or higher than 5% of the simulated F-values generated by 1000 Monte Carlo permutations (i.e., P < 0.05). Analyses were performed using CANOCO 4.5 software (Ter Braak and Smilauer, 2002).
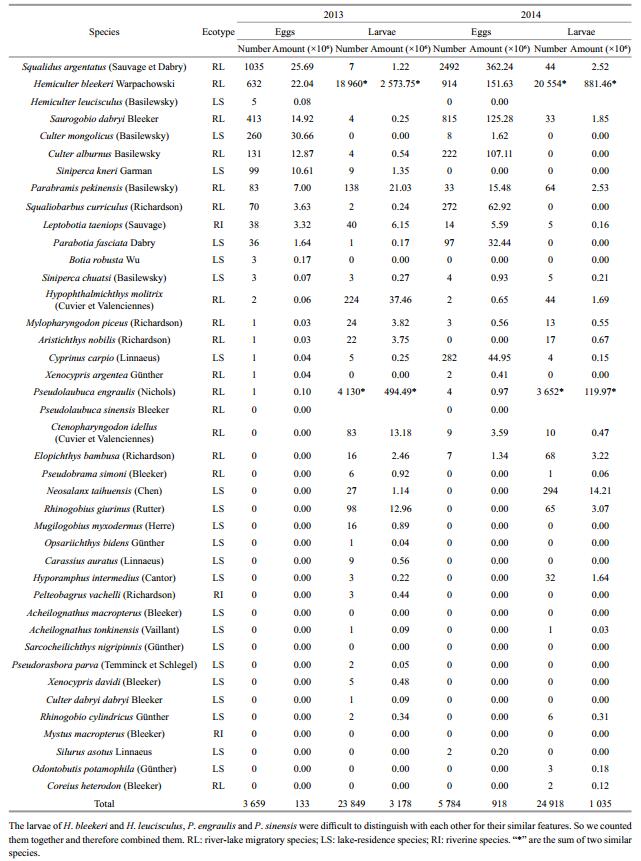
3 RESULT 3.1 Species composition and total amount of ichthyoplanktonA total of forty-one species of eggs and larvae belonging to five orders and nine families were identified on 150 sampling days in two years. Among these samples, 16 were RL, such as the four major Chinese carps (Mylopharyngodon piceus, Ctenopharyngodon idella, Hypophthalmichthys molitrix, Aristichthys nobilis) (Table 1). All the egg samples belonged to 21 species, including 13 RL. Meanwhile, the larvae specimens belong to 34 species, including 15 RL.
|
A total of 130 million eggs and 3 180 million larvae were estimated drifting through the sampling section from May 23 to July 24 in 2013. The eggs comprised an estimated 25.7 million Squalidus argentatus, 14.9 million Saurogobio dabryi, 22.0 million Hemiculter bleekeri, 12.9 million Culter alburnus, and 30.7 million Culter mongolicus. All the larvae mainly consisted of Hemiculter (2 570 million) and Pseudolaubuca (490 million). The estimated total amount of eggs and larvae for the RL reached 3 210 million, accounting for 97.0% of all the ichthyoplankton captured (Table 1).
From May 1 to July 26 in 2014, an estimated 1 060 million eggs and 1 040 million larvae drifted through the sampling section. The eggs comprised an estimated 362 million S. argentatus, 125 million S. dabryi, 152 million H. bleekeri, 107 million C. alburnus, 62.9 million Squaliobarbus curriculus and 44.9 million Cyprinus carpio. Hemiculter (880 million) and Pseudolaubuca (120 million) dominated the larvae. A total of 1 850 million eggs and larvae were recorded for RL, accounting for 94.6% of all the ichthyoplankton captured (Table 1).
3.2 Temporal variations in recruitment 3.2.1 Temporal patterns of eggsA total of 8 443 fish eggs were collected on 150 sampling days. In the Songzi channel, seasonal changes in drift density followed a multimodal pattern (Fig. 2). Five peaks of drift eggs measured by density were evident from late May to July in 2013. The first peak appeared on June 2 dominated by S. argentatus and S. dabryi. The second peak, which was the highest of the five (506 ind./1 000 m3), appeared on June 8 and was dominated by H. bleekeri, S. argentatus, and S. dabryi. The following three peaks appeared in midJune (June 16), late June (June 27) and mid-July (July 11). The third peak was dominated by species similar to those of the second peak, whereas H. bleekeri dominated the fourth peak. The fifth peak was dominated by C. alburnus, C. mongolicus and Siniperca kneri.

|
| Fig.2 Daily variation in egg drift density (column) and water level (solid line) in the Songzi channel from May to July in 2013 and 2014 |
In 2014, five peaks of egg density were observed, and they appeared in mid-May (May 13), late May (May 21), early June (June 3), late June (June 21), and early July (July 4). H. bleekeri dominated the first, third, and fourth peaks, S. argentatus dominated all four peaks, and S. dabryi dominated the first three peaks. However, in the fifth peak, the highest peak, the dominant species changed, and only C. alburnus remained abundant.
Species composition showed distinctly varied from May to July in both years. Several species, such as S. argentatus, S. dabryi and H. bleekeri, spawned from early May to late July and dominated in May. On the other hand, other species, such as C. alburnus, Parabramis pekinensis, S. curriculus and Parabotia fasciata, started spawning from late June (Fig. 3).

|
| Fig.3 Monthly density percent (%) of the dominant groups in eggs collected in the Songzi channel from May to July in 2013 and 2014 |
A total of 48 767 fish larvae were collected on 150 sampling days. Larvae mainly appeared in late July, corresponding to the water level and (or) water discharge (Fig. 4). In 2013, larvae drift density was the highest on July 14th and dominated by Hemiculter and Pseudolaubuca. In 2014, three peaks of drift density appeared on July 12 (2 424 ind./1 000 m3), 20 (1 403 ind./1 000 m3), and 24 (3 561 ind./1 000 m3). The peaks were all dominated by Hemiculter and Pseudolaubuca.

|
| Fig.4 Daily variation in larva drift density (column) and water level (solid line) in the Songzi channel from May to July in 2013 and 2014 |
By screening the environmental variables whose inflation factors reached more than 20 and variables with P > 0.05 using Monte Carlo permutation tests, we identified two variables, flow and water temperature, relating with egg drift density. With CCA, we plotted species in an ordination figure with environmental variables representing as vectors. The direction and length of vectors represented the influence of environmental variables on ichthyoplankton drift density. We estimated the correlation between ichthyoplankton drift density of different species and environmental variables by vertical projection of species on the vectors. The projections in the direction of the vectors indicated positive correlation between ichthyoplankton drift density and environmental variables against the direction implied negative correlation. Results showed that spawning of S. curriculus, Cobitidae, P. pekinensis, C. alburnus, Siniperca and C. mongolicus were strongly influenced by flooding (represented by water discharge or water level) and water temperature, whereas H. bleekeri, S. argentatus and S. dabryi spawned with little requirement for the two environmental factors (Fig. 5).

|
| Fig.5 Biplot of species and environmental variables in 2013 and 2014 H. ble: Hemiculter bleekeri; S. arg: Squalidus argentatus; S. dab: Saurogobio dabryi; C. mon: Culter mongolicus; C. alb: Culter alburnus; P. pek: Parabramis pekinensis; S. cur: Squaliobarbus curriculus. |
Floodplain channels are the main sources and paths of rivers supplying water to floodplains. Some fish species may use floodplain channel systems as corridors to spawning grounds or nursery grounds during high water flow periods (Copp, 1989), which are linked with increased diversity and yield of fishes in riverine systems (Burgess et al., 2013). Bénech and Peñáz (1995) reported that the considerable portion of recruitment in fish populations in the floodplain originates from the main river, with larvae and young juveniles entering the floodplain for nursery and growth in the Central Delta of the Niger River. Sixteen species of RL were sampled in the present study, these fish species will play important roles in fish diversity in the Dongting Lake. According to studies, these sixteen fish species represent 23.5% of the species in the west Dongting Lake (a part of the Dongting Lake directly connecting with the Songzi channel) (Zhu et al., 2014). In the present study, huge amuont of ichthyoplankton (3 350 million in 2013 and 2 100 million in 2014), especially RL, flow into the Dongting Lake and play important roles in fish recruitment in the Dongting Lake.
The river-lake migratory fish of Dongting Lake partly originate from its own tributaries and mostly from the Changjiang mainstream (Zhou et al., 1986). River-lake migratory fishes represent 23.6% of the fishes in the Dongting Lake (Tang and Qian, 1979). In 1963, the production of river-lake migratory fishes accounted for 32% of the fishery yield in the Dongting Lake (Li et al., 2013a). The river-lake migratory fishes from the Changjiang River also play vital roles in the fish resources in the Dongting Lake.
In recent decades, reduction in fish diversity and abundance occurred in numerous Changjiang lakes due to the construction of sluice gates leading to riverlake disconnection, environmental pollution, and overfishing. For example, in Honghu Lake, fish species decreased from 74 in the 1960s to 57 in the 1990s (Chen and Xu, 1995) and in Donghu Lake, fish species dropped from 67 in the 1960s to 38 in the 1990s (Huang and Xie, 1996). We suspect that without ichthyoplankton recruitment from the Changjiang mainstream, fish diversity and abundance in the Dongting Lake will also dramatically decrease.
4.2 Environmental factors and fish resource conservationWhile water level and (or) water flow fluctuated, ichthyoplankton recruitment also fluctuated over time during the sampling period. In the present study, evident temporal patterns were observed for ichthyoplankton recruitment. Some species bred earlier in the season, for example S. argentatus, S. dabryi, and H. bleekeri while some species bred relatively late, for example, C. alburnus, C. mongolicus, and P. pekinensis. Spawning of most fish species was significantly correlated with flooding (Figs. 2 and 5). For example, rheophilic species (i.e. S. curriculus) and some eurytopics (i.e. C. mongolicus, C. alburnus, P. pekinensis and Siniperca) show strong requirement for flood pulse for spawning (Fig. 5). These findings agree with the results of other researchers (Yi and Liang, 1964; Yi et al., 1988a; Qiu et al., 2002; Wang et al., 2008). Flooding plays an important role in spawning of RL such as the four major Chinese carps, Coreius guichenoti, C. heterodon, and Leptobotia elongata (Yu et al., 1984; Yi et al., 1988a; Cao et al., 2007). As the typical river-lake migratory fishes, the four major Chinese carps are also important commercial fish species and have been studied extensively in the Changjiang Basin (Wu et al., 1988; Yi et al., 1988a; Qiu et al., 2002; Tan et al., 2007; Jiang et al., 2010). Li et al. (2013b) reported that the most favorable flood condition for spawning of the four major Chinese carps involves the diurnal increase in water level and water discharge to values higher than 0.55 m/d and 2 100 m3/(s∙d), respectively. Meanwhile, the drift density of most fish species was also suggested to correspond with water discharge (Jiang et al., 2010). Particularly, Zhou et al. (1986) considered that the percentage of ichthyoplankton of the four major Chinese carps flowing through the three inlet channels is approximately equal to the percentage of water discharge. Therefore, water discharge is very crucial to ichthyoplankton drift.
Given the heavy human activities, the water discharge flowing through the three inlet channels has sharply decreased (Li et al., 2009; Fang et al., 2014; Hu et al., 2014). In 1994–2002, and 2003–2011, the mean discharge was 616m8 m3/s and 500m8 m3/s, 32.1% and 44.9% less than long term average one (911m8 m3) (Hu et al., 2014), respectively. This decrease in discharge not only caused the shrinking and siltation of the channel, but also hindered the recruitment of ichthyoplankton to the Dongting Lake (Zhou et al., 1986).
To prevent the further demise of the channels, irrigation measures were proposed for the three channels, and a sluice gate was suggested for the Songzi channel (Wang et al., 2013). For the conservation of fish resource and increasing ichthyoplankton recruitment, we suggested to irrigate the channel to increase water discharge as this action may increase the transport of ichthyoplankton from the Changjiang mainstream to the Dongting Lake. However, we advised against building the sluice gate because such structure may prevent ichthyoplankton flow into the lake.
5 CONCLUSIONAs a result of the construction of sluice gate, which led to river-lake disconnection, environmental pollution, and overfishing, fish diversity in a number of Changjiang lakes sharply decreased. Dongting Lake, as one of the only three lakes remaining connected with the Changjiang mainstream, maintains high fish diversity and production. Our research shows the still high abundances of ichthyoplankton recruitment to the Dongting Lake from the Changjiang River. Owing to heavy human activities, water discarge flowing through the three inlet channels, which flow into the Dongting Lake, has sharply decreased, which will cause a decrease in ichthyoplankton recruitment. Therefore, we suggest the irrigation of the channel to increase water discharge.
6 DATA AVAILABILITY STATEMENTThe datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Bénech V, Peñáz M. 1995. An outline on lateral fish migrations within the Central Delta of the Niger River, Mali. Hydrobiologia, 303(1-3): 149-157.
DOI:10.1007/BF00034052 |
Burgess O T, Pine W E, Walsh S J. 2013. Importance of floodplain connectivity to fish populations in the Apalachicola River, Florida. River Research and Applications, 29(6): 718-733.
DOI:10.1002/rra.2567 |
Cao W X, Chang J B, Qiao Y, Duan Z H. 2007. Fish Resources of Early Life History Stages in Yangtze River. China Water Power Press, Beijing.
(in Chinese)
|
Cao W X. 2011. Current situation and protection countermeasures of fish resources in the Yangtze River. Jiangxi Fishery Science and Technology, 2: 1-3.
(in Chinese) |
Chang J B, Cao W X. 1999. Fishery significance of the rivercommunicating lakes and strategies for the management of fish resources. Resources and Environment in the Yangtze Basin, 8(2): 153-157.
(in Chinese with English abstract) |
Chen D Q, Duan X B, Liu S P, Shi W G, Wang B. 2002. On the dynamics of fishery resources of the Yangtze River and its management. Acta Hydrobiologica Sinica, 26(6): 685-690.
(in Chinese with English abstract) |
Chen Y Y, Xu Y X. 1995. Hydrobiology and Resources Exploitation in Honghu Lake. Science Press, Beijing.
(in Chinese)
|
Copp G H. 1989. The habitat diversity and fish reproductive function of floodplain ecosystems. Environmental Biology of Fishes, 26(1): 1-27.
DOI:10.1007/BF00002472 |
Dou H S, Jiang J H. 2000. Dongting Lake. University of Science and Technology of China Press, Hefei.
(in Chinese)
|
Duan X B, Liu S P, Huang M G, Qiu S L, Li Z H, Wang K, Chen D Q. 2009. Changes in abundance of larvae of the four domestic chinese carps in the middle reach of the Yangtze River, China, before and after closing of the three gorges dam. Environmental Biology of Fishes, 86(1): 13-22.
DOI:10.1007/s10641-009-9498-z |
Fang C M, Chen W J, Zhou H M, Zhang Y P, Fu P F, He G, Wu B, Wang S. 2016. Fish resources and its utilization suggestions of Lake Poyang. Jiangsu Agricultural Science, 44(9): 233-242, 243.
(in Chinese) |
Fang C M, Hu C H, Chen X J. 2014. Impacts of Three Georges Reservoir's operation on outflow of the three outlets of Jingjiang River and Dongting Lake. Journal of Hydraulic Engineering, 5(1): 36-41.
(in Chinese with English abstract) |
Fang J Y, Wang Z H, Zhao S Q, Li Y K, Tang Z Y, Yu D, Ni L Y, Liu H Z, Xie P, Da L J, Li Z Q, Zheng C Y. 2006. Biodiversity changes in the lakes of the Central Yangtze. Frontiers in Ecology and the Environment, 4(7): 369-377.
DOI:10.1890/1540-9295(2006)004[0369:BCITLO]2.0.CO;2 |
Han Q W, Zhou S H. 1999. Characteristics and fluvial processes of diffluent rivers from Changjiang River to Dongting Lake. Journal of Yangtze River Scientific Research Institute, 16(5): 5-8.
(in Chinese with English abstract) |
Hu G W, Mao D H, Li Z Z, Zeng Y, Feng C. 2014. Characteristics and driving factors of runoff and sediment changes fluxes into the Dongting Lake from three outlets in Jingjiang river during past 60 years. Journal of Natural Resources, 29(1): 129-141.
(in Chinese with English abstract) |
Huang G T, Xie P. 1996. Changes in the structure of fish community with the analysis on the possible reasons in Lake Donghu, Wuhan. Acta Hydrobiologica Sinica, 20(S1): 38-46.
(in Chinese with English abstract) |
Humphries P, Serafini L G, King A J. 2002. River regulation and fish larvae: Variation through space and time. Freshwater Biology, 47(7): 1307-1331.
DOI:10.1046/j.1365-2427.2002.00871.x |
Jiang W, Liu H Z, Duan Z H, Cao W X. 2010. Seasonal variation in drifting eggs and larvae in the upper Yangtze, China. Zoological Science, 27(5): 402-209.
DOI:10.2108/zsj.27.402 |
Kottelat M, Whitten T. 1996. Freshwater biodiversity in Asia with special reference to fish. In: World Bank Technical Paper, No. 343. The World Bank, Washington DC. p.17-21, https: //doi.org/10.1596/0-8213-3808-0.
|
Li J Q, Wang D L, Ding D M. 2013a. Research progress of fish resources in Dongting lake. Journal of Anhui Agricultural Science, 41(9): 3898-3900.
(in Chinese with English abstract) |
Li M Z, Duan Z H, Gao X, Cao W X, Liu H Z. 2016. Impact of the Three Gorges Dam on reproduction of four major Chinese carps species in the middle reaches of the Changjiang River. Chinese Journal of Oceanology and Limnology, 34(5): 885-893.
DOI:10.1007/s00343-016-4303-2 |
Li M Z, Gao X, Yang S R, Duan Z H, Cao W X, Liu H Z. 2013b. Effects of environmental factors on natural reproduction of the four major Chinese carps in the Yangtze River, China. Zoological Science, 30(4): 296-303.
DOI:10.2108/zsj.30.296 |
Li T T, Shen Y. 2005. A study on biodiversity conversation in Dongting Lake area. Territory & Natural Resources Study, (1): 38-40.
(in Chinese with English abstract) |
Li Y T, Guo X H, Tang J W, Sun Z H. 2009. Changes on runoff diversion from Jingjiang Reach of the Yangtze River to Dongting Lake after the operation of Three Gorges Reservoir. Journal of Basic Science and Engineering, 17(1): 21-31.
(in Chinese with English abstract) |
Liang Z S, Zhou C S, Huang H N. 1981. Composition and seasonal changes of fishes in Lake Wuhu connected with Changjiang River. Oceanoligia et Limnologia Sinica, 12(5): 468-478.
(in Chinese with English abstract) |
Liao F C, He W, Huang X R, Jing Q L, He X C. 2002. Studies on present situation and change trend of Dongting Lake fishery resources and enviroment. Acta Hydrobiologica Sinica, 26(6): 623-627.
(in Chinese with English abstract) |
Liao F C, He X C, He W, Wang H W, Xu D P. 2006. Status and protective regulation countermeasure in fishery resources and its environment of Dongting Lake. Journal of Yueyang Vocational Technical College, 21(6): 32-37.
(in Chinese with English abstract) |
Martens K, Segers H. 2009. Endemism in aquatic ecosystems. In: Encyclopedia of Inland Waters. Academic Press, Oxford. p.423-430.
|
Mu H X, Li M Z, Liu H Z, Cao W X. 2014. Analysis of fish eggs and larvae flowing into the three gorges reservoir on the Yangtze River, China. Fisheries Science, 80(3): 505-515.
DOI:10.1007/s12562-014-0729-7 |
Olson D M, Dinerstein E. 2002. The global 200: priority ecoregions for global conservation. Annals of the Missouri Botanical Garden, 89(2): 199-224.
DOI:10.2307/3298564 |
Qiu S L, Liu S P, Huang M G, Chen D Q, Duan X B. 2002. Monitoring of spawning sites of four major Chinese carps in the middle sector of Yangtze River. Acta Hydrobiologica Sinica, 26(6): 716-718.
(in Chinese) |
Ru H J, Liu X Q, Huang X R, Ning Y Z, Wang H Z. 2008. Diversity of fish species and its spatio-temporal variations in Lake Dongting, a large Yangtze-connected lake. Journal of Lake Sciences, 20(1): 93-99.
(in Chinese with English abstract) DOI:10.18307/2008.0114 |
Sun S L, Shen J. 2015. Research on evolution and regulation of the Songzi river. Journal of Water Resources Research, 4(6): 559-566.
(in Chinese with English abstract) DOI:10.12677/JWRR.2015.46070 |
Tan X C, Li X H, Tao J P, Li J, Wang C, Luo J R, Lai Z N. 2007. Spatial and temporal distribution of larval rusources of fishes in Xijiang River. Freshwater Fisheries, 37(4): 37-40.
(in Chinese with English abstract) |
Tang J H, Qian M Q. 1979. Fish fauna in Dongting Lake. Inland Fisheries, (10): 24-32.
(in Chinese) |
Ter Braak C J F, Prentice I C. 1988. A theory of gradient analysis. Advances in Ecological Research, 18: 271-317.
DOI:10.1016/S0065-2504(08)60183-X |
Ter Braak C J, Smilauer P. 2002. Canoco reference manual and canodraw for windows user's guide: Software for canonical community ordination (Version 4. 5). Microcomputer Power, New York.
|
Tockner K, Stanford J A. 2002. Riverine flood plains: Present state and future trends. Environmental Conservation, 29(3): 308-330.
DOI:10.1017/S037689290200022X |
Wang C P, Yu Q H, Zhang L M. 2013. Preliminary discussion on relevant issues of sluice construction at Songzi Entrance on Jinjiang River. Yangtze River, 44(24): 20-22.
(in Chinese with English abstract) |
Wang S Y, Liao W G, Chen D Q, Duan X B, Peng Q D, Wang K, Li C. 2008. Analysis of ecohydrological characteristics of the four Chinese farmed carps spawning grounds in the middle reach of the Yangtze River. Resources and Environment in the Yangtze Basin, 17(6): 298-798.
(in Chinese with English abstract) |
Wu G X, Liu L H, Wang Z L, Cao W X, Wang Z X. 1988. Investigation on natural reproduction of Ctenopharyngodon idellus in Jinshajiang of upper Yangtze River. Freshwater Fisheries, (1): 3-6.
(in Chinese) |
Xie P. 2017. Ecological impacts of three Gorges dam on Lakes Dongting and Poyang. Resources and Environment in the Yangtze Basin, 26(10): 1.
(in Chinese with English abstract) |
Yi B L, Liang Z S, Yu ZY, Lin R D, He M J. 1988b. A comparative study of the early development of grass carp, black carp, silver carp and big head of the Yangtze River. In: Yi B, Yu Z, Liang Z eds. Gezhouba Water Control Project and Four Domestic Fishes in Yangtze River. Hubei Science and Technology Press, Wuhan, China. p.111-135. (in Chinese with English abstract)
|
Yi B L, Liang Z S. 1964. Natural conditions of the spawning grounds of the domestic fishes in Changjiang River and essential external factor for spawning. Acta Hydrobiologica Sinica, 5: 1-15.
(in Chinese with English abstract) |
Yi B L, Yu Z T, Liang Z S, Shen S J, Xu Y G, Chen J X, He M J, Liu Y L, Hu Y Z, Deng Z L, Huang S W, Sun J Y, Liu R J, Xiang Y. 1988a. The distribution, natural conditions and breeding production of the spawning groups of four famous freshwater fishes on the main stream on the Yangtze River. In: Yi B L, Yu Z T, Liang Z S eds. Gezhouba Water Control Project and Four Famous Fishes in Yangtze River. Hubei Science and Technology Press, Wuhan, China. p.1-46. (in Chinese with English abstract)
|
Yu Z T, Liang Z S, Yi B L. 1984. The early development of Coreius heterodon and Coreius guichenoti. Acta Hydrobiologica Sinica, 8(4): 371-388.
(in Chinese with English abstract) |
Zhou C S, Xiang Y, Sun J Y, Tan D Q, Yao C C, Duan Z H. 1986. Prediction of effects of the Three Gorge project on fish resources in lakes along the middle Changjiang River and study of possible countermeasure. In: The Leading Group of the Three Gorge Project on Ecology and Environment Research Project, Chinese Academy of Science ed. The Symposium on Impacts of the Three Gorge Project on Ecology and Environment and Possible Countermeasures. Science Press, Beijing. p.21-29.
|
Zhu Y, Lü C, Hu H J, Wang Z S, Jia Y F, He M Y, Huang X J, Lei G C. 2014. Changes in fish community structure in West Dongting Lake after the operation of the Three Gorges Dam. Journal of Lake Sciences, 26(6): 844-852.
(in Chinese with English abstract) DOI:10.18307/2014.0605 |