 2020, Vol. 38
2020, Vol. 38Institute of Oceanology, Chinese Academy of Sciences
Article Information
- CHENG Huihui, LIU Wenbin, YUAN Xiangyang, JIA Erteng, ZHANG Dingdong, JIANG Guangzhen
- Effects of dietary cottonseed meal protein hydrolysate on growth, antioxidants and immunity of Chinese mitten crab Eriocheir Sinensis
- Journal of Oceanology and Limnology, 38(3): 869-882
- http://dx.doi.org/10.1007/s00343-019-9152-3
Article History
- Received Jun. 8, 2019
- accepted in principle Aug. 14, 2019
- accepted for publication Sep. 19, 2019




Chinese mitten crab Eriocheir sinensis is one of the most economically valuable freshwater crabs in Asia (Rudnick et al., 2003). It was reported that world aquaculture output of Chinese mitten crab reached 796 622 tons in 2014 (Pauly and Zeller, 2017). However, with the development of aquaculture practice, a variety of diseases caused by viruses (Zhang and Bonami, 2012) and bacteria (Wang and Gu, 2002) may emerge and affect the production of Chinese mitten crab. When bacteria or viruses infect crabs, it would affect the health of crabs and consequently their growth, thus result in a burst of death (Fu et al., 2017). Therefore, it is important to strengthen the resistance of crabs. In the past few years, to reduce the risk of bacterial and viral infections in crabs, many researches carried out studies to enhance the antioxidant capacity and immune response of crabs, for example, supplying nutrient suitably of feed (Sun et al., 2013b; Wei et al., 2016), and adding small bioactive peptides (Guo et al., 2017) and immune-enhancers moderately (Sun et al., 2013a; Hong et al., 2017; Jia et al., 2017). However, most of the researchers concentrated on juvenile Eriocheir sinensis, and research on adult ones remains poor. As crustaceans, it is necessary to work more to enhance or regulate the immunological response of adult crab in order to avoid alteration of the immune system (Sánchez et al., 2001).
Plant protein as an ingredient of aquaculture feed is less cost compared with animal protein, especially fish meal. However, the utilization of plant proteins (e.g., cottonseed meal, rapeseed meal) is limited due to their antinutritional factors such as gossypol (Jiang et al., 2018). These antinutritional factors can cause negative effect on growth and health of aquatic animals (Figueiredo-Silva et al., 2015; Jiang et al., 2018). Plant protein hydrolysate is produced using processes such as hydrolytic enzymes (Colla et al., 2014; Muranova et al., 2017). Studies has proven that these hydrolysates are ideal nutrient sources, they could increase weight gain, improve immunity and antioxidant capability of aquatic animals when added within a suitable range (Gibbs et al., 2004; Kotzamanis et al., 2007; Möller et al., 2008; Gui et al., 2010; Yimit et al., 2012; Song et al., 2014).
A previous study by Liu (2005) suggested that cottonseed meal protein hydrolysate (CPH) had the best effect on the growth and immunity of fish compared with those hydrolysates of rapeseed meal, soybean meal, and peanut meal. Thus, we chose cottonseed meal (CM) to produce CPH. The proportions of small molecule peptide and free amino acid consisting in CPH were significantly improved compared with CM. The peptide content of CPH was 13.4% (180–1 983 Da), whereas CM was only 1.9% (180–1 983 Da) (Gui et al., 2010). CPH can improve feed ingredient palatability as an attractant of Chinese mitten crab (Cheng et al., 2019), and boost immunity of Alogynogenetic crucian carp (Liu, 2005); it can also improve the activities of serum lysozyme and alkaline phosphatase in the intestines of Cyprinus carpio var. jian (Xia et al., 2012). However, presently, there is no available information on the effects of dietary CPH supplementation on immunity of Chinese mitten crab. Thus, the main aim of our study is to explore the effects of dietary CPH at different levels on growth, antioxidant capability and immune response as well as the mRNA expression levels of immune-related genes of Chinese mitten crab.
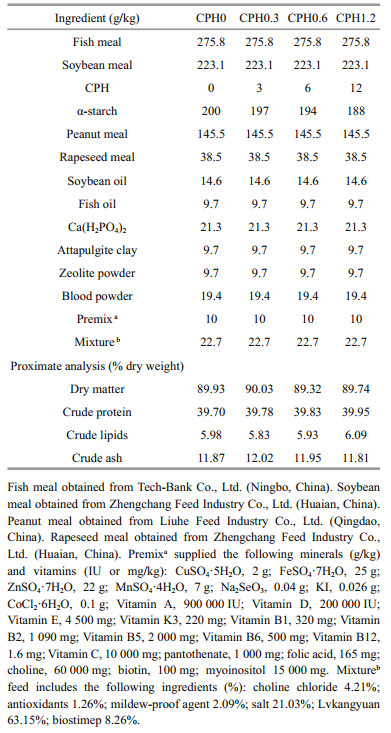
2 MATERIAL AND METHOD 2.1 Experimental ingredients and dietsCPH was produced by the method of Xia et al. (2012). Cottonseed meal protein was hydrolysated by AS1.398 protease in water bath (5 h, 45℃, pH 7.0). Four different CPH dosages were added to the basic diets to make the experimental diets: 0% (CPH0 group), 0.3% (CPH0.3 group), 0.6% (CPH0.6 group), and 1.2% (CPH1.2 group). Ingredients and proximate composition of the experimental diets are shown in Table 1. Fish meal, soybean meal, peanut meal, and rapeseed meal were supplemented as protein sources. Soybean oil and fish oil were supplemented as lipid sources, and α-starch was supplemented as carbohydrate source. All diets were prepared in laboratory. Feed ingredients were ground into fine powder, and then they mixed with oil and water to be dough. After that, the dough was extruded through a single-screw meat grinder extruder. These feeds were air-dried and then were cut to appropriate sizes. Last, they were stored in sealed bags at -20℃.
Crabs were obtained from a local feeding farm in Pukou, Jiangsu Province, China. In the beginning, crabs were fed a basal diet twice a day to adopt experimental conditions for 2 weeks. Then, 160 crabs (initial weight: 51.32±0.08 g, mean±SEM) were randomly assigned into 20 cement pools (width×length×height, 2.0 m×2.0 m×0.8 m) at a density of 8 crabs per cement pool, five replicates was contained in each group. Crabs were fed twice daily (07:00 am and 05:00 pm) for 60 days. The number of deaths crabs was recorded daily. During the experimental period, water temperature ranged from 23 to 30℃, pH fluctuated between 7.2 and 7.4, and dissolved oxygen was maintained above 5.0 mg/L.
2.3 Sample collectionAt the end of the feeding trial, one crab from each cement pool was sampled at random for analysis of the whole body composition. Three crabs from per replicate were anesthetized by an MS-222 (tricaine methanesulfonate, Sigma, USA) at a concentration of 150 mg/L. After that, they were weighed, respectively. Hemolymph was obtained using syringes from each crab's second last pair of walking legs 1:1 with precooling anticoagulant solution (citrate, 26 mmol/L; glucose, 100 mmol/L; citric acid, 30 mmol/L; NaCl, 450 mmol/L; EDTA, 15 mmol/L; pH=7.2) (Li et al., 2014), with that, hemolymph was centrifuged immediately at 3 500 r/min, 4℃ for 20 min. The supernatant serum was sucked up in the centrifuge tube, and then they were stored at -80℃ for subsequent analysis. After that, every crab was dissected, and then obtained hepatopancreas were put in a liquid nitrogen container. Finally, they were stored at -80℃ for subsequent analysis.
2.4 Perfusion experimentAnother group of crabs (mean weight 85 g) were acclimated for 2 weeks with commercial diet at 28℃ in one 200-L tank with circulating water system. Crabs were starved for 24 h before perfusion.
Six crabs were randomly collected and anesthetized by an MS-222 (tricaine methanesulfonate, Sigma, USA) at a concentration of 150 mg/L, and after that, they were dissected, obtained hepatopancreas and intestine, which were used for time 0. Then, 30 crabs were divided into two groups and weighted respectively. Crabs in two groups were perfused intragastrically with either saline solution (10 mL per kg crab body weight) or CPH solution (10 mL per kg crab body weight), which a saline solution (0.9%) containing 5 mg CPH per mL. The two groups were labeled as Sham group and CPH group. After that, crabs were transferred quickly to 10 tanks (100 L each) at a density of three crabs per tank. After perfusion, crabs in each group were sampled at 3, 6, 12, 18, and 24 h, respectively. One tank was used for each sampling time so as to minimize the stress during sampling. Hepatopancreas and intestine were sampled and then were kept at -80℃ for subsequent analysis of gene expression.
2.5 Parameters measurement and methods 2.5.1 Crabs growth performanceThe body weight of each crab was determined at the start and end of the feeding experiment. In addition, the specific growth rate (SGR) and survival rate (SR) were determined using the following formula:
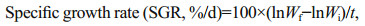
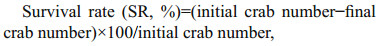
where Wf represented final body weight (g), Wi represented initial body weight (g), t represented the total experimental days.
2.5.2 Proximate analysis of the experimental diets and body compositionThe diets and the crabs were analyzed to determine their proximate composition according to the procedures detailed by the AOAC (1984). The moisture contents of diets and crabs were estimated by using drying oven to a constant weight; ash was determined by combustion at 550℃ for 4–6 h; crude protein was determined using an Auto Kjeldahl System (1030-Auto-analyzer, Tecator, Hoganas, Sweden) by using the micro-Kjeldahl method; crude lipid was determined using a Soxtec System HT (Soxtec System HT6, Tecator, Hoganas, Sweden) by solvent extraction; the energy levels of diets and crabs were determined using a Bomb Calorimeter (Parr 1281, Parr Instrument Company, Moline, IL, USA).
2.5.3 Analysis of antioxidant capabilitiesHemolymph and hepatopancreas were used to estimate the antioxidant capability of crabs. The hepatopancreas was weighed accurately and homogenized in an ice bath with nine volumes (v/w) of chilled saline in a tissue homogenizer, and then the extract was centrifuged at 9 000 r/min at 4℃ for 15 min. Finally, all the supernatant was then stored at -20℃ for subsequent analysis.
Total superoxide dismutase (t-SOD) activity in hemolymph was detected according to the method described by Wang and Chen (2005). Superoxide dismutase (SOD) activity in hepatopancreas was detected using the Ransod Kit (Randox, Crumlin, UK) by the ability of inhibiting superoxide radical-dependent reactions. Catalase (CAT) activity was measured according the method described by Lygren et al. (1999). Malondialdehyde (MDA) content was estimated using the thiobarbituric acid test (Satoh, 1978).
2.5.4 Immune parameters in hemolymphHemolymph was used to measure the activities of lysozyme (LZM), acid phosphatase (ACP) and alkaline phosphatase (ALP). The activity of LZM was assayed according to LZM kit protocol at 530 nm according to Yuan et al. (2007). The activities of ACP and ALP were measured by the disodium phenyl phosphate hydrate method (Chen et al., 2016).
2.5.5 GPT and GOT activities in hemolymphThe activities of glutamic-oxaloacetic transaminase (GOT) and glutamic-pyruvic transaminase (GPT) in hemolymph and their standard curves were measured by the spectrophotometer at 520 nm, then the activities of GPT and GOT were calculated according to the standard curves (Bergmeyer et al., 1978).
2.5.6 Analysis of gene expressionTotal RNA was extracted using RNAiso Plus (TaKaRa, Dalian, China). The total RNA purity was detected using NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE). The RNA samples were used for the subsequent analysis only when the A260/A280 ratio between 1.8 and 2.0. The reverse transcription-PCR reaction of PrimeScriptTM RT Master Mix (TaKaRa, Dalian, China) was: 15 min at 37℃, 5 s at 85℃, and after that, 4℃ thereafter. The obtained cDNA was kept at -20℃.
Real-time quantitative PCR (RT-qPCR) primers for immune-related genes are presented in Table 2. RT-qPCR was performed on a real-time PCR detection system (ABI USA-7500) using a SYBR Premix Ex Taq II kit (TaKaRa, Dalian, China). The RT-qPCR reactions were performed in a final 20-μL volume, including 10 μL SYBR Premix Ex TaqⅡ (2x), 0.4 μL primer (10 μm), 0.4 μL ROX, 6.8 μL DEPC water, as well as 2 μL of cDNA template. The reactions were initially denatured at 95℃ for 10 min and then 40 cycles at 95℃ for 15 s, followed by annealing at 60℃ for 34 s. The relative mRNA levels were calculated by the 2-△△Ct method (Livak and Schmittgen, 2001), then, β-Actin was acted as the endogenous control to normalize the mRNA expression of the immune-related genes.

|
The data on feeding trial was subjected to One-way analysis of variance (One-way ANOVA) followed by Turkey's test using the SPSS version 22.0 software (SPSS Inc., Chicago, IL, USA). The data regarding gene expression levels in the CPH perfusion and Sham perfusion were analyzed by Two-way ANOVA for significant differences among treatment means based on sampling time, perfusion type and their interaction. If the interaction were significant (P < 0.05) differences, each factor was further analyzed separately by One-way ANOVA. Specifically, data among the different treatments at each sampling time was analyzed by One-way ANOVA. Data among different sampling time within each treatment was analyzed by t-test. The data are presented as mean±S.E.M.
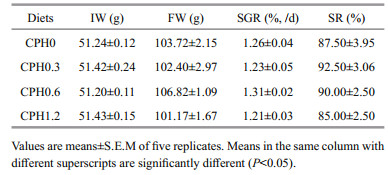
3 RESULT 3.1 Growth performanceAs can be seen from Table 3, no difference (P > 0.05) were seen in FW, SGR and SR in crabs fed with CPH diet compared to the CPH0 group.
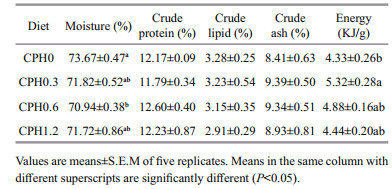
As shown in Table 4, crude protein, lipid, and ash content all shows no differences (P > 0.05) among all the groups. However, moisture content was significantly reduced (P < 0.05) in crabs fed with CPH0.6 diet compared to the CPH0 group, but no difference (P > 0.05) was observed among other supplemented diet. Similarly, energy content was significantly higher (P < 0.05) in crabs fed with CPH0.3 diet compared to the CPH0 group, but no noticeable difference (P > 0.05) was observed among others.
|
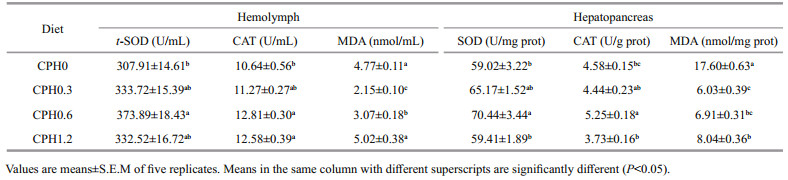
As shown in Table 5, SOD and CAT activities in hemolymph and hemolymph of crabs fed with CPH0.6 diet were significantly (P < 0.05) higher than that of the CPH0 group whereas the CPH0.3 and CPH0.6 groups was not different significantly (P > 0.05). MDA content in hemolymph of crabs fed with CPH0.3 diet was significantly lower than those of the other groups in terms of dietary CPH levels (P < 0.05). MDA content in hepatopancreas of crabs fed with CPH0.3 diet was significantly (P < 0.05) lower than that of the CPH0 group whereas the CPH0.3 and CPH0.6 groups was not different significantly (P > 0.05).
As shown in Fig. 1: LZM activity in hemolymph of crabs fed with CPH0.3 diet was significantly (P < 0.05) higher than those of the other groups in terms of dietary CPH levels; ALP had the same tendency with LZM, and it was significantly (P < 0.05) higher than those of the other groups in terms of dietary CPH levels; ACP activity in hemolymph of crabs fed with CPH0.3 diet was significantly (P < 0.05) higher than the CPH0 group whereas the CPH0.3 and CPH0.6 groups was not different significantly (P > 0.05). GPT activity in hemolymph of crabs fed with CPH0.3 diet was significantly (P < 0.05) lower than that of the CPH0 group, although there was no significant difference between CPH0.3 and CPH0.6 groups (P > 0.05). There was no significant difference in GOT activity among all groups (P > 0.05).

|
| Fig.1 LZM (a), ALP (b), ACP(c), GPT (d), and GOT (e) activities in hemolymph of crabs fed diet containing different levels of CPH Each datum represents the mean of five replicates. Bars assigned with different superscripts are significantly different (P < 0.05). |
As shown in Fig. 2, the relative expression level of Toll1 of crabs fed with CPH0.3 diet was significantly (P < 0.05) higher than that of the CPH0 group and CPH1.2 group. The relative expression level of Toll2 of crabs fed with CPH0.3 diet was significantly (P < 0.05) higher than those of the other groups in terms of dietary CPH levels. The relative expression levels of MyD88 and LITAF of crabs fed with CPH0.3 diet were significantly (P < 0.05) higher than those of the other groups in terms of dietary CPH levels. Similarly, the relative expression level of ILF-2 of crabs fed with CPH0.3 diet was significantly (P < 0.05) higher than those of the other groups in terms of dietary CPH levels. The relative expression levels of SOCS2 of crabs fed with CPH0.3 diet was significantly (P < 0.05) lower than that of the CPH0 group.

|
| Fig.2 Relative expression levels of Toll1 (a), Toll2 (b), MyD88 (c), LITAF (d), ILF-2 (e), and SOCS2 (f) in hepatopancreas of crabs fed diet containing different levels of CPH Each datum represents the mean of five replicates. Bars assigned with different superscripts are significantly different (P < 0.05). |
As shown in Figs. 3 & 4, after CPH perfusion, the relative expression levels of Toll1, Toll2, MyD88, LITAF, ILF-2 in intestine and hepatopancreas were all significantly (P < 0.01) affected by perfusion type, time and their interaction; SOCS2 expression level was significantly (P < 0.001) affected by time and the interaction of time and perfusion type. The CPH treatment resulted in the expression levels of Toll1 at times 3 h, 18 h and Toll2 at times 18 h, 24 h in intestine significantly (P < 0.05) increase after perfusion. However, they declined significantly (P < 0.01) at other times. After the CPH treatment, Toll1 and Toll2 expression in hepatopancreas and MyD88 expression in intestine and hepatopancreas decreased significantly (P < 0.05) with the lowest value at 6 h. LITAF expression in intestine and hepatopancreas decreased significantly (P < 0.05) with the lowest value at 12 h. The expression level of ILF-2 in intestine at time 3 h and in hepatopancreas at times 6 h decreased significantly (P < 0.05) to the lowest value. After that, they increased gradually until basal level was reached compared to Sham group. The expression level of SOCS2 in intestine at time 3 h and in hepatopancreas at times 18 h increased significantly (P < 0.05) to the highest value. The expression levels of all these genes did not difference at all times after sham treatment.

|
| Fig.3 Relative expressions of Toll1 (a), Toll2 (b), MyD88 (c), LITAF (d), ILF-2 (e) and SOCS2 (f) in intestine of crabs after CPH perfusion Data are referred to the value (relative units, RU) obtained in the corresponding tissue of the sham treatment at 0 h. Each data represents the mean of three replicates. Significant differences (P < 0.05) among sampling times within each treatment are indicated by different letters (lower-case letters for CPH, capital letters for sham). * represents a significant difference (P < 0.05) between the two groups at each sampling time. *P < 0.05, **P < 0.01, ***P < 0.001; ns: not significant. |

|
| Fig.4 Relative expressions of Toll1 (a), Toll2 (b), MyD88 (c), LITAF (d), ILF-2 (e) and SOCS2 (f) in hepatopancreas of crabs after CPH perfusion Data are referred to the value (relative units, RU) obtained in the corresponding tissue of the sham treatment at 0 h. Each data represents the mean of three replicates. Significant differences (P < 0.05) among sampling times within each treatment are indicated by different letters (lower-case letters for CPH, capital letters for sham). * represents a significant difference (P < 0.05) between the two groups at each sampling time. *P < 0.05, **P < 0.01, ***P < 0.001; ns, not significant. |
The growth of animal is affected by feed utilization rate and the nutrients deposition of animal is greatly influenced by the nutritional composition of feed (Sheridan and Mommsen, 1991; Duan and Plisetskaya, 1993). It is well known that crabs grow quickly only after molting (Hartnoll, 2001; He, 2005). The molting period is influenced by temperature, nutrition, and breeding environment (Chen et al., 1995; Saucedo et al., 2004; He, 2005). The above could explain why there was no significant difference in SGR and SR of crabs. However, Gui et al. (2010) found that supplement of dietary CPH could increase the body weight of crucian carp (Carassius auratus gibelio), it may be that crustacean growth only occurs during molting (He, 2005).
It has been reported that the differences in dietary nutrients would possibly cause the differences in body composition (Hansen et al., 2007). In the present study, dietary supplement with 0.6% CPH could decrease the moisture content of crabs, whereas the opposite was true for crude protein content, indicating that CPH may promote the accumulation of organic matter. This result was possible that the lower molecular weight, small peptides, and free amino acids in CPH could promote protein synthesis by activating TOR signaling pathway, this needs further study. Our study also found that the energy content of crabs fed CPH0.3 diet increased significantly, it might be that absorbed dietary amino acids and peptides were processed to increase energy content of total dry matters in crabs. Overall, our study suggested that CPH might have no impact on the puberty molt, but it could promote the deposition of nutrients of crabs.
The body cells produce reactive oxygen species (ROS) under normal physiological conditions (Zhang et al., 2013). However, the imbalance of generation and elimination of ROS will result in oxidative stress (Shen et al., 2010; Wu et al., 2013), then, SOD and CAT would be treated as the first line of the antioxidant defense system against ROS produced in the organism (Li et al., 2007). In the study, enhancing SOD and CAT activities and reducing MDA content were seen in crabs fed diets containing CPH, suggesting that the CPH could improve the antioxidant capacity of crabs.Previous study has reported that small peptides could stimulate and activate the antioxidant capacity of animal body and increase the activities of antioxidant enzymes (Chen et al., 1998; Moure et al., 2006). So, the results of our study were probably related to the rich concentration of small peptides in CPH. Similarly, Wang et al. (2017) studied that fermented cottonseed meal could increase serum antioxidant enzymes activities in broilers. Fermented mushroom bran hydrolysate replacement fishmeal could significantly decrease the content of MDA in serum of the allogynogenetic crucian carp (Zhang et al., 2017). Adding soy protein hydrolysates in diets enhanced SOD activity and decreased MDA content in serum of juvenile starry flounder (Song et al., 2014). However, in the present study, high dietary CPH inclusion level might provoke oxidative stress. It may be that high level CPH could stimulate the ROS release largely resulting in the imbalance of generation and elimination. Similarly, it was reported that prawn fed with a high fermented soybean meal level diet might induce oxidative stress (Matozzo et al., 2011). Overall, our study suggested that appropriate supplement CPH could promote the antioxidant capability of crabs.
LZM is regarded as an important non-specific immune response indicator of crustaceans and has bactericidal activity in many studies (Yano et al., 1996; Fu et al., 2017). ACP and ALP also play an important role in crustacean non-specific immune defence (Matozzo et al., 2011). In the present study, the activities of LZM, ALP, and ACP of crabs in hemolymph were significantly increased when dietary supplement with 0.3% CPH. This indicated that an optimum administration of CPH might improve the immune function of crabs. It might be that the small molecule peptide consisting in CPH had an important role in boosting immunity. It was similar with our results that the appropriate amount of CPH could improve the non-specific immunity of the Cyprinus carpio var. jian (Xia et al., 2012). Previous similar studies showed that fermented cottonseed meal could enhance immune function of yellow-feathered broilers (Tang et al., 2012), and increase serum immunoglobulin level of broilers (Wang et al., 2017). Another similar study shown that fish fed with soy protein hydrolysates diet could increase LZM activity significantly (Song et al., 2014).
GPT and GOT can be widely found in the mitochondria of many tissues and organs, particularly in the liver cells (Zheng and Pu, 1997). In the present study, we found that the activity of GPT in the hemolymph of crabs declined as dietary CPH was 0.3%. It indicated that appropriate supplementation of CPH could protect the hepatopancreas from damage, implying that CPH had a protective mechanism to the hepatopancreas of crabs. It may be that CPH can inhibit harmful bacteria and viruses infections. Nevertheless, high concentration of CPH significantly increased the activity of GPT, it might be that the hepatic cells were injured, and then the GPT transported into blood, resulting the increase of GPT activities in hemolymph (Wang et al., 2005). The reason might be that high CPH level might be harmful to hepatic cells by enhancing amino acid metabolism and producing more metabolic wastes. A similar study result appeared in juvenile starry flounder after feeding with soy protein hydrolysates (Song et al., 2014). Therefore, our study indicated that supplementation of moderate CPH was useful to protect hepatopancreas of crabs. This may be that the CPH could enhance immune function of crabs. Moreover, the SR of crabs in different groups was consistent with these result, this further suggested that dietary CPH improved the resistance of crab by the CPH-induced immune responses.
Next, we examined the relative expressions of Toll1, Toll2, MyD88, LITAF, ILF-2, and SOCS2 in the hepatopancreas of crabs. Tolls play an important role in eliminating extracellular invaders, and they can be regarded as the bridge linking the innate and adaptive immunity (Lin et al., 2012). LITAF is a key transcription factor in downstream of Toll signaling pathway, and it is contacted with the transcriptional regulation of cytokines dependent on MyD88 (Li et al., 2014). SOCS2 could be induced by bacteria challenge and played a key role in the immune defense response of crabs (Zhang et al., 2010). In the present study, we found that the relative expression levels of Toll1 and Toll2 were significantly increased after feeding CPH0.3 diet. It suggested that Toll1 andToll2 might be potential receptor of CPH to regulate the downstream transcription factors. It was known that Tolls might participate in antifungal and antibacterial immune responses, and it was beneficial to the ability of crabs to start a suitable immune response against different pathogen-associated molecular patterns (Yu et al., 2013). The relative expression level of MyD88 increased significantly after feeding CPH0.3 diet. This could be conjectured that the moderate supplement of CPH could promote the activation of the downstream signal factors by MyD88 in crabs. It was reported that MyD88 play a key role in activating the immune system of crabs to resist the bacterial infections (Huang et al., 2014). Dietary supplement with 0.3% CPH could increase the expression level of LITAF of crabs, the result indicated that feeding CPH0.3 diet might have a stimulating role to the downstream cytokines of Toll signaling pathway. The relative expression level of LITAF was consistent with MyD88, suggesting that the immune function of CPH on Toll signaling pathway may be shown by affecting its downstream cytokines through MyD88. The relative expression level of ILF-2 increased when dietary CPH level was 0.3%, suggesting that ILF2 might induce all kinds of immune-related gene expression (Wang et al., 2006). However, the expression of SOCS2 declined after feeding CPH0.3 diet. It may be that SOCS2 was only induced after bacteria challenge. These results further suggested that dietary CPH could improve the immunity of crabs.
After that, we performed a perfusion experiment to explore further the effect of short-term CPH stimulant on immune gene expression of crabs. In intestine, the highest expression level of Toll1 and SOCS2 were obtained at time 3 h, indicating that the immune function of Toll1 and SOCS2 were firstly activated by CPH, in hepatopancreas, the highest expression level of SOCS2 was obtained at time 18 h, suggesting that the immune response in intestine was faster than in hepatopancreas, a probably reason is that the CPH first acted on the intestine. Moreover, the expression level of SOCS2 was affected by time and the interaction of time and perfusion type, which suggested that SOCS2 could be induced by CPH and probably played an important role in immune defense responses of crabs. Evidence suggests that the transcripts encoding SOCS proteins in cells are usually at low or undetectable levels, but they can induced rapidly by cytokines and pathogen components widely in M. musculus (Zhang et al., 2010). Our results were similar to those discoveries. However, other genes expression levels declined firstly and then increased gradually, suggesting that these genes could not rapidly respond to CPH stimulation. The expression level of these genes decreased at CPH stimulation might indicate that this period belonged to the convalescence of crab after CPH stimulation. It took least 6 h for these genes to reanimate and then produced effect in innate immune responses. This is similar to the findings that the expression level of SpToll in hemocytes of Scylla paramamosain dealt with poly I:C. or V. parahemolyticus (Lin et al., 2012) and the expression level of FcToll of Fenneropenaeus chinensis in the early period after exposure to bacteria (Yang et al., 2008). Another reason we speculated that these genes may have additional functional roles. It was reported that certain genes were only expressed when the epithelial cells in 'mature zone' of the organ was damaged (Hauton et al., 2006). However, further studies should be performed to explore the reason at a deeper level.
5 CONCLUSIONIn summary, this experiment showed that dietary supplementation of CPH has beneficial effects on the accumulation of organic matter, antioxidant capacity, and immune response that might via the activation of immune-related genes such as Tolls and MyD88; the appropriate supplement dosage of CPH for Chinese mitten crab could be 0.3%–0.6%. However, the precise mechanism requires further study.
6 DATA AVAILABILITY STATEMENTThe data that support the findings of this study are available from the corresponding author upon reasonable request.
Association of Official Analytical Chemists (AOAC). 1984.Official Methods of Analysis. 14th edn. AOAO, Washington, 1018p.
|
Bergmeyer H U, Scheibe P, Wahlefeld A W. 1978. Optimization of methods for aspartate aminotransferase and alanine aminotransferase. Clinical Chemistry, 24(1): 58-73.
DOI:10.1093/clinchem/24.1.58 |
Chen H M, Muramoto K, Yamauchi F, Fujimoto K, Nokihara K. 1998. Antioxidative properties of histidine-containing peptides designed from peptide fragments found in the digests of a soybean protein. Journal of Agricultural and Food Chemistry, 46(1): 49-53.
DOI:10.1021/jf970649w |
Chen J C, Lin M N, Ting Y Y, Lin J N. 1995. Survival, haemolymph osmolality and tissue water of Penaeus chinensis juveniles acclimated to different salinity and temperature levels. Comparative Biochemistry and Physiology Part A:Physiology, 110(3): 253-258.
DOI:10.1016/0300-9629(94)00164-O |
Chen Y L, Chen L Q, Qin J G, Ding Z L, Li M, Jiang H B, Sun S M, Kong Y Q, Li E C. 2016. Growth and immune response of Chinese mitten crab (Eriocheir sinensis) fed diets containing different lipid sources. Aquaculture Research, 47(6): 1 984-1 995.
DOI:10.1111/are.12654 |
Cheng H H, Jiang G Z, Zheng X C, Xu C Y, Sun C X, Zhang D D, Liu W B. 2019. Effects of five attractants on Chinese mitten crab, Eriocher sinensis. Acta Hydrobiologica Sinica, 43(2): 395-401.
(in Chinese with English abstract) |
Colla G, Rouphael Y, Canaguier R, Svecova E, Cardarelli M. 2014. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Frontiers in Plant Science, 5: 448.
|
Duan C, Plisetskaya E M. 1993. Nutritional regulation of insulin-like growth factor-I mRNA expression in salmon tissues. Journal of Endocrinology, 139(2): 243-252.
|
Figueiredo-Silva C, Lemme A, Sangsue D, Kiriratnikom S. 2015. Effect of DL-methionine supplementation on the success of almost total replacement of fish meal with soybean meal in diets for hybrid tilapia (Oreochromis niloticus×Oreochromis mossambicus). Aquaculture Nutrition, 21(2): 234-241.
DOI:10.1111/anu.12150 |
Fu L L, Zhou G, Pan J L, Li Y H, Lu Q P, Zhou J, Li X G. 2017. Effects of Astragalus polysaccharides on antioxidant abilities and non-specific immune responses of Chinese mitten crab, Eriocheir sinensis. Aquaculture International, 25(3): 1 333-1 343.
DOI:10.1007/s10499-017-0117-2 |
Gibbs B F, Zougman A, Masse R, Mulligan C. 2004. Production and characterization of bioactive peptides from soy hydrolysate and soy-fermented food. Food Research International, 37(2): 123-131.
DOI:10.1016/j.foodres.2003.09.010 |
Gui D, Liu W B, Shao X P, Xu W N. 2010. Effects of different dietary levels of cottonseed meal protein hydrolysate on growth, digestibility, body composition and serum biochemical indices in crucian carp (Carassius auratus gibelio). Animal Feed Science and Technology, 156(3-4): 112-120.
DOI:10.1016/j.anifeedsci.2010.01.012 |
Guo Z L, Qiao X, Cheng R M, Shi N N, Wang A L, Feng T T, Chen Y, Zhang F, Yu H N, Wang Y P. 2017. As-CATH4 and 5, two vertebrate-derived natural host defense peptides, enhance the immuno-resistance efficiency against bacterial infections in Chinese mitten crab, Eriocheir sinensis. Fish & Shellfish Immunology, 71: 202-209.
|
Hansen A C, Rosenlund G, Karlsen Ø, Koppe W, Hemre G I. 2007. Total replacement of fish meal with plant proteins in diets for Atlantic cod (Gadus morhua L. ) I - effects on growth and protein retention. Aquaculture, 272(1-4): 599-611.
|
Hartnoll R G. 2001. Growth in Crustacea-twenty years on. Hydrobiologia, 449(1-3): 111-122.
|
Hauton C, Brockton V, Smith V J. 2006. Cloning of a crustinlike, single whey-acidic-domain, antibacterial peptide from the haemocytes of the european lobster, Homarus gammarus, and its response to infection with bacteria. Molecular Immunology, 43(9): 1 490-1 496.
DOI:10.1016/j.molimm.2005.07.029 |
He J. 2005. Population growth characteristics of the mitten crab in ecological aquaculture ponds. Reservoir Fisheries, 25(6): 10-11, 28.
(in Chinese with English abstract) |
Hong Y H, Yang X Z, Yan G W, Huang Y, Zuo F, Shen Y X, Ding Y, Cheng Y X. 2017. Effects of glyphosate on immune responses and haemocyte DNA damage of Chinese mitten crab, Eriocheir sinensis. Fish & Shellfish Immunology, 71: 19-27.
|
Huang Y, Chen Y H, Wang Z, Wang W, Ren Q. 2014. Novel myeloid differentiation factor 88, EsMyD88, exhibits EsTube-binding activity in Chinese mitten crab Eriocheir sinensis. Developmental & Comparative Immunology, 47(2): 298-308.
|
Jia E T, Li Z Q, Xue Y F, Jiang G Z, Li X F, Liu W B, Zhang D D. 2017. Effects of dietary fructooligosaccharide on the growth, antioxidants, immunity and disease resistance of Chinese mitten crab. Aquaculture, 481: 154-161.
DOI:10.1016/j.aquaculture.2017.08.033 |
Jiang H B, Chen L Q, Qin J G. 2018. Fishmeal replacement by soybean, rapeseed and cottonseed meals in hybrid sturgeon Acipenser baerii ♀×Acipenser schrenckii ♂. Aquaculture Nutrition, 24(4): 1 369-1 377.
DOI:10.1111/anu.12674 |
Kotzamanis Y P, Gisbert E, Gatesoupe F J, Infante J Z, Cahu C. 2007. Effects of different dietary levels of fish protein hydrolysates on growth, digestive enzymes, gut microbiota, and resistance to Vibrio anguillarum in European sea bass (Dicentrarchus labrax) larvae. Comparative Biochemistry and Physiology Part A:Molecular & Integrative Physiology, 147(1): 205-214.
|
Li S, Jia Z R, Li X J, Geng X Y, Sun J S. 2014. Identification and expression analysis of lipopolysaccharide-induced TNF-alpha factor gene in Chinese mitten crab Eriocheir sinensis. Fish & Shellfish Immunology, 38(1): 190-195.
|
Li X M, Ma Y L, Liu X J. 2007. Effect of the Lycium barbarum polysaccharides on age-related oxidative stress in aged mice. Journal of Ethnopharmacology, 111(3): 504-511.
DOI:10.1016/j.jep.2006.12.024 |
Lin Z Y, Qiao J, Zhang Y L, Guo L L, Huang H, Yan F, Li Y Y, Wang X Y. 2012. Cloning and characterisation of the SpToll gene from green mud crab, Scylla paramamosain. Developmental & Comparative Immunology, 37(1): 164-175.
DOI:10.1016/j.dci.2011.09.003 |
Liu W B. 2005. Effect of Plant Protein Hydrolysates on the Growth Development of Alogynogenetic crucian and BioActive Valve Analysis of Plant Protein Hydrolysates. Nanjing Agricultural University, Nanjing.
(in Chinese with English abstract)
|
Livak K J, Schmittgen T D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25(4): 402-408.
|
Lygren B, Hamre K, Waagbø R. 1999. Effects of dietary proand antioxidants on some protective mechanisms and health parameters in atlantic salmon. Journal of Aquatic Animal Health, 11(3): 211-221.
DOI:10.1577/1548-8667(1999)011<0211:EODPAA>2.0.CO;2 |
Matozzo V, Gallo C, Monari M, Marin M G. 2011. Cellular and biochemical parameters in the crab Carcinus aestuarii after experimentally-induced stress:effects of bacterial injection, leg ablation and bacterial injection/leg ablation combination. Journal of Experimental Marine Biology and Ecology, 398(1-2): 18-25.
DOI:10.1016/j.jembe.2010.11.014 |
Möller N P, Scholz-Ahrens K E, Roos N, Schrezenmeir J. 2008. Bioactive peptides and proteins from foods:indication for health effects. European Journal of Nutrition, 47(4): 171-182.
DOI:10.1007/s00394-008-0710-2 |
Moure A, Domínguez H, Parajó J C. 2006. Antioxidant properties of ultrafiltration-recovered soy protein fractions from industrial effluents and their hydrolysates. Process Biochemistry, 41(2): 447-456.
|
Muranova T A, Zinchenko D V, Kononova S V, Belova N A, Miroshnikov A I. 2017. Plant protein hydrolysates as fish fry feed in aquaculture. Hydrolysis of rapeseed proteins by an enzyme complex from king crab hepatopancreas.Applied Biochemistry and Microbiology, 53(6): 680-687.
|
Pauly D, Zeller D. 2017. Comments on FAOs State of World Fisheries and Aquaculture (SOFIA 2016). Marine Policy, 77: 176-181.
DOI:10.1016/j.marpol.2017.01.006 |
Rudnick D A, Hieb K, Grimmer K F, Resh V H. 2003. Patterns and processes of biological invasion:the Chinese mitten crab in San Francisco Bay. Basic and Applied Ecology, 4(3): 249-262.
DOI:10.1078/1439-1791-00152 |
Sánchez A, Pascual C, Sánchez A, Vargas-Albores F, Moullac G L, Rosas C. 2001. Hemolymph metabolic variables and immune response in Litopenaeus setiferus adult males:the effect of acclimation. Aquaculture, 198(1-2): 13-28.
DOI:10.1016/S0044-8486(00)00576-7 |
Satoh K. 1978. Serum lipid peroxide in cerebrovascular disorders determined by a new colorimetric method. Clinica Chimica Acta, 90(1): 37-43.
DOI:10.1016/0009-8981(78)90081-5 |
Saucedo P E, Ocampo L, Monteforte M, Bervera H. 2004. Effect of temperature on oxygen consumption and ammonia excretion in the calafia mother-of-pearl oyster, Pinctada mazatlanica (Hanley, 1856). Aquaculture, 229(1-4): 377-387.
DOI:10.1016/S0044-8486(03)00327-2 |
Shen W Y, Fu L L, Li W F, Zhu Y R. 2010. Effect of dietary supplementation with Bacillus subtilis on the growth, performance, immune response and antioxidant activities of the shrimp (Litopenaeus vannamei). Aquaculture Research, 41(11): 1 691-1 698.
DOI:10.1111/j.1365-2109.2010.02554.x |
Sheridan M A, Mommsen T P. 1991. Effects of nutritional state on in vivo lipid and carbohydrate metabolism of coho salmon, Oncorhynchus kisutch. General and Comparative Endocrinology, 81(3): 473-483.
|
Song Z D, Li H Y, Wang J Y, Li P Y, Sun Y Z, Zhang L M. 2014. Effects of fishmeal replacement with soy protein hydrolysates on growth performance, blood biochemistry, gastrointestinal digestion and muscle composition of juvenile starry flounder(Platichthys stellatus). Aquaculture, 426-427: 96-104.
DOI:10.1016/j.aquaculture.2014.01.002 |
Sun R, Yue F, Qiu L M, Zhang Y, Wang L L, Zhou Z, Zhang H, Yi QL, Song L S. 2013a. The CpG ODNs enriched diets enhance the immuno-protection efficiency and growth rate of Chinese mitten crab, Eriocheir sinensis. Fish & Shellfish Immunology, 35(1): 154-160.
|
Sun S M, Qin J G, Yu N, Ge X P, Jiang H B, Chen L Q. 2013b. Effect of dietary copper on the growth performance, nonspecific immunity and resistance to Aeromonas hydrophila of juvenile Chinese mitten crab, Eriocheir sinensis. Fish & Shellfish Immunology, 34(5): 1 195-1 201.
|
Tang J W, Sun H, Yao X H, Wu Y F, Wang X, Feng J. 2012. Effects of replacement of soybean meal by fermented cottonseed meal on growth performance, serum biochemical parameters and immune function of yellowfeathered broilers. Asian-Australasian Journal of Animal Sciences, 25(3): 393-400.
DOI:10.5713/ajas.2011.11381 |
Wang H J, Xiang L X, Shao J Z, Jia S. 2006. Molecular cloning, characterization and expression analysis of an IL-21 homologue from Tetraodon nigroviridis. Cytokine, 35(3-4): 126-134.
DOI:10.1016/j.cyto.2006.07.010 |
Wang S H, Chen J C. 2005. The protective effect of chitin and chitosan against Vibrio alginolyticus in white shrimp Litopenaeus vannamei. Fish & Shellfish Immunology, 19(3): 191-204.
|
Wang W, Gu Z F. 2002. Rickettsia-like organism associated with tremor disease and mortality of the Chinese mitten crab Eriocheir sinensis. Diseases of Aquatic Organisms, 48(2): 149-153.
|
Wang Y W, Deng Q Q, Song D, Wang W W, Zhou H, Wang L, Li A K. 2017. Effects of fermented cottonseed meal on growth performance, serum biochemical parameters, immune functions, antioxidative abilities, and cecal microflora in broilers. Food and Agricultural Immunology, 28(4): 725-738.
DOI:10.1080/09540105.2017.1311308 |
Wang Y, Xiong L, Yang K J, Wu Z, Sheng X M, Tang H F, Liu X P. 2005. Effect of beta-cypermethrin on GPT and GOT activities of crucian serum. Agricultural Science & Technology, 6(1): 20-23.
|
Wei J J, Zhang F, Tian W J, Kong Y Q, Li Q, Yu N, Du Z Y, Wu Q Q, Qin J G, Chen L Q. 2016. Effects of dietary folic acid on growth, antioxidant capacity, non-specific immune response and disease resistance of juvenile Chinese mitten crab Eriocheir sinensis (Milne-Edwards, 1853). Aquaculture Nutrition, 22(3): 567-574.
DOI:10.1111/anu.12275 |
Wu J Q, Kosten T R, Zhang X Y. 2013. Free radicals, antioxidant defense systems, and schizophrenia. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 46(1): 200-206.
|
Xia W, Liu W B, Qiao Q S, Li G F, Zhang Y J. 2012. Effects of cottonseed meal hydrolysate on growth performance and biochemical indices of Cyprinus carpio var. Jian. Freshwater Fisheries, 42(1): 46-51.
(in Chinese with English abstract) |
Yang C J, Zhang J Q, Li F H, Ma H M, Zhang Q L, Priya T A J, Zhang X J, Xiang J H. 2008. A Toll receptor from Chinese shrimp Fenneropenaeus chinensis is responsive to Vibrio anguillarum infection. Fish & Shellfish Immunology, 24(5): 564-574.
|
Yang J L, Wang L L, Huang M M, Wang L L, Gai Y C, Qiu L M, Zhang H, Song L S. 2011. An interleukin-2 enhancer binding factor 2 homolog involved in immune response from Chinese mitten crab Eriocheir sinensis. Fish & Shellfish Immunology, 30(6): 1 303-1 309.
|
Yano T. 1996. The nonspecific immune system:humoral defense. Fish Physiology, 15: 105-157.
DOI:10.1016/S1546-5098(08)60273-3 |
Yimit D, Hoxur P, Amat N, Uchikawa K, Yamaguchi N. 2012. Effects of soybean peptide on immune function, brain function, and neurochemistry in healthy volunteers. Nutrition, 28(2): 154-159.
DOI:10.1016/j.nut.2011.05.008 |
Yu A Q, Jin X K, Guo X N, Li S, Wu M H, Li W W, Wang Q. 2013. Two novel Toll genes (EsToll1 and EsToll2) from Eriocheir sinensis are differentially induced by lipopolysaccharide, peptidoglycan and zymosan. Fish & Shellfish Immunology, 35(4): 1 282-1 292.
|
Yuan C T, Li D M, Chen W, Sun F F, Wu G H, Gong Y, Tang J Q, Shen M F, Han X D. 2007. Administration of a herbal immunoregulation mixture enhances some immune parameters in carp (Cyprinus carpio). Fish Physiology and Biochemistry, 33(2): 93-101.
DOI:10.1007/s10695-006-9120-7 |
Zhang C N, Li X F, Xu W N, Jiang G Z, Lu K L, Wang L N, Liu W B. 2013. Combined effects of dietary fructooligosaccharide and Bacillus licheniformis on innate immunity, antioxidant capability and disease resistance of triangular bream (Megalobrama terminalis). Fish & Shellfish Immunology, 35(5): 1 380-1 386.
|
Zhang D S, Zhang Y P, Liu B, Jiang Y, Zhou Q L, Wang J, Wang H L, Xie J, Kuang Q. 2017. Effect of replacing fish meal with fermented mushroom bran hydrolysate on the growth, digestive enzyme activity, and antioxidant capacity of allogynogenetic crucian carp (Carassius auratus gibelio). Turkish Journal of Fisheries and Aquatic Sciences, 17(5): 1 039-1 048.
|
Zhang S, Bonami J R. 2012. Isolation and partial characterization of a new reovirus in the Chinese mitten crab, Eriocheir sinensis H Milne Edwards. Journal of Fish Diseases, 35(10): 733-739.
DOI:10.1111/j.1365-2761.2012.01398.x |
Zhang Y, Zhao J M, Zhang H, Gai Y C, Wang L L, Li F M, Yang J L, Qiu L M, Song L S. 2010. The involvement of suppressors of cytokine signaling 2 (SOCS2) in immune defense responses of Chinese mitten crab Eriocheir sinensis. Developmental & Comparative Immunology, 34(1): 42-48.
|
Zhao D, Song L, Liu R, Liang Z, Wang L, Sun M. 2016. The immunosuppressive effects of continuous CpG ODNs stimulation in Chinese mitten crab, Eriocheir sinensis. Invertebrate Survival Journal, 13: 34-43.
|
Zheng Y H, Pu F Y. 1997. Effect of mercury on transaminase activities of tissues in C parpio & C auratus. Journal of Southwest Agricultural University, 19(1): 41-45.
(in Chinese with English abstract) |